1. 식후 및 공복상태에서의 대사 조절
식사후에 소화관으로부터 혈액으로 유입되는 상태인 식후 흡수상태(absorptive state)와 신체에 저장되어 있는 영양소를 에너지원으로 사용하는 공복상태(postabsorptive state)로 나누어 설명
일반적으로 식사 후부터 흡수가 완료되기 까지 4시간 소요되며, 사람들은 하루 3식 식습관을 가지고 있으므로 일반적으로 늦은 오전시간, 늦은 오후 시간 및 밤 시간동안 인체는 공복상태가 된다. 사람이 24시간 이상 음식을 먹지 않은 상태를 단식상태(fasting state)라고 한다. 식후 흡수상태에서는 흡수된 영양소 중 일부는 필요한 에너지를 공급하는데 사용되고, 나머지는 에너지 저장장소에서 저장되었다가 다음에 오는 공복상태에서 사용된다.
식후 흡수상태
그림에 식후 흡수상태일때 각 조직에서 일어나는 대사과정을 요약
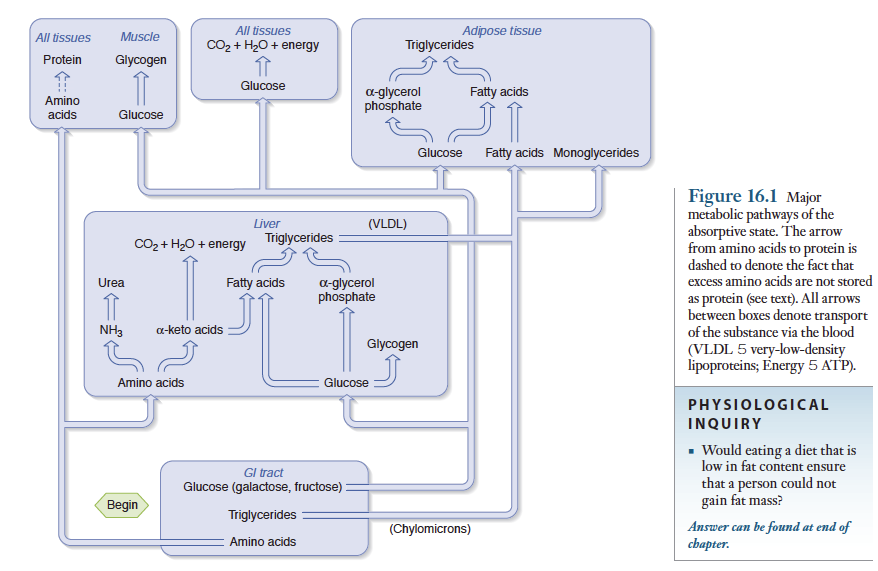
3대 영양소(탄수화물, 지방, 단백질) 중 탄수화물이 주된 에너지원
15장에서 설명했듯이 탄수화물과 단백질은 단당류와 아미노산으로 분해되어 흡수된 후 소화관에 분포되어 있는 모세혈관으로 흡수. 소화관에서 흡수된 영양소가 풍부한 혈액은 간문맥을 통해 간으로 직접 이동. 간에서 나온 혈액은 심장을 통해 신체 각 부위로 전달되기 때문에 간은 혈액이 신체 각 부위에 이동되기 전에 혈액의 영양소 조성을 변화시키는 역할을 함.
탄수화물, 아미노산과는 달리 지방은 소장에서 흡수된 후 유미입자 상태로 림프계로 들어감. 이 림프액은 간을 거치지 않고 체순환계의 정맥으로 합류되므로, 지방이 신체 각 부위로 이동되기 전에 간조직이 지방성분을 변화시키지는 못함.
참고) 다음에 심장순환 탐구때 간으로 이어지는 혈관, 혈액순환을 전체적으로 그려내야...
흡수된 탄수화물
소화관에서 흡수된 탄수화물 중 일부는 갈락토오스와 과당이지만, 이들 당류들은 간에서 포도당으로 전환된 후 포도당과 똑같은 과정에 의해 대사되므로 흡수된 탄수화물을 포도당으로 간주.
포도당은 식후 몇시간 동안 인체의 주된 에너지원으로 작용. 흡수된 포도당은 세포속으로 들어가 CO2와 물로 분해되면서 에너지와 ATP를 공급. 골격근은 체질량의 상당부분을 차지하기 때문에 포도당을 에너지원으로 소비하는 주된 조직이며, 휴식상태에서도골격근은 포도당을 주로 이용. 골격근은 식후에 포도당을 분해하여 에너지원으로 이용할뿐 아니라 일부 포도당을 글리코겐으로 전환하여 근육에 저장.
지방세포도 포도당을 에너지원으로 이용하지만, 식후에 지방세포에서 일어나는 주된 대사과정은 포도당이 트리글리세리드로 전환되는 것임. 포도당으로부터 알파-글리세롤 인산과 지방산이 생성되며, 지방세포는 이 두 물질을 결합하여 트리글리세리드를 합성. 흡수된 탄수화물 중 대부분은 간으로 들어감. 식후에 간으로 유입된 포도당은 골격근에서와 같이 글리코겐으로 저장되거나, 지방조직에서와 같이 알파-글리세롤 인산과 지방산으로 전환되어 트리글리세리드 합성에 이용됨. 간에서 포도당으로부터 합성된 지방은 일부 간에 저장되기도 하지만, 대부분 특정 단백질과 결합하여 초저밀도 지방단백질(Very low density lipoprotein, VLDL)이라하는 지방단백질을 형성. 이 지방단백질은 간세포로부터 분비되어 혈액으로 들어감. VLDL은 단백질보다는 지방을 많이 함유하는데, 지방의 밀도는 단백질보다 낮으므로 이 지방단백질을 VLDL이라 함. 간세포에서 VLDL이 합성되는 과정은 소장 점막세포에서유미입자가 형성되는 과정과 유사함.
VLDL은 크기가 큰 지방단백질이므로 일단 혈류로 들어가면 모세혈관벽을 벽을 쉽게 통과하지 못함. 그 대신 VLDL 내의 트리글리세리드는 모세혈관 내피세포의 표면에 있는 지방단백질리파아제(lipoprotein lipase)에 의해 단당류(즉 글리세롤)와 지방산으로 분해됨. 지방단백질리파아제는 특히 지방조직의 모세혈관에 풍부함. 이때 지방조직의 모세혈관에서 VLDL로부터 생성된 지방산은 모세혈관벽을 통해 지방세포 속으로 확산되어 들어감. 지방세포내에서 지방산은 포도당의 대사산물인 알파-글리세롤 인산과 결합하여 트리글리세리드를 다시 형성함. 따라서 VLDL에 있는 대부분의 지방산은 그 기원이 간조직에서 포도당으로부터 합성된 VLDL의 글리세리드로부터 온 것이며, 최종적으로 지방조직에 트리글리세리드 형태로 저장됨. 지방단백질리파아제의 작용에 의해 지방 조직의 모세혈관에 있는 형성된 혈액내 모노글리세리드는 간으로 되돌아가서 대사됨.
요약하면, 식후에 흡수된 포도당의 주된 대사경로는 에너지원으로 이용되거나 간과 골격근에 글리코겐으로 저장되거나 지방조직에지방으로 저장됨.
흡수된 지질들
흡수된 유미입자는 림프액으로 들어가서 혈액과 합류함. 혈중의 유미입자가 대사되는 생화학적 과정은 VLDL이 대사되는 과정과 매우 유사함. 지방조직을 관류하는 모세혈관 내벽의 지방단백질리파아제의 작용으로 혈장내 유미입자로부터 지방산이 유리됨. 유리된 지방산은 지방세포로 들어가 그곳에서 포도당대사에 의해 생성된 알파-글리세롤 인산과 결합하여 글리세리드로 전환된 후 저장됨.
이와같이 지방세포에서 트리글리세리드가 합성되기 위해서는 포도당이 필수적임. 지방세포는 글리세롤을 인산화하는 효소가 없으므로 글리세롤이나 다른 지방대사산물로부터 알파-글리세롤 인산을 생성할 수 없으며, 유일하게 탄수화물 중간대사과정에서 생성됨. 알파-글리세롤 인산과 달리 지방조직의 트리글리세리드 합성을 위해 다음과 같이 세가지 공급원으로부터 지방산이 올 수 있음.1) 지방조직에 유입된 포도당으로부터 합성된 지방산 2) 간에 유입된 포도당이 지방산으로 전환된 후 VLDL 트리글리세리드 상태로 지방조직으로 운반된 후 생성된 지방산 3) 식사로부터 흡수된 지방산이 유미입자 트리글리세리드 상태로 지방조직에 운반되어 생성된 지방산임. 위의 과정 중 2)와 3)은 지방단백질리파아제의 작용에 의해 혈중 트리글리세리드로부터 유리되어 지방조직으로 들어감.
유미입자에 들어있는 지방 중 콜레스테롤은 에너지원으로 이용되지 않으며, 주된 역할은 세포막의 구성성분일 뿐만 아니라 담즙산염, 스테로이드 호르몬의 전구물질로 작용하므로 체내에서 중요한 역할을 함. 그러나 불행히도 혈중 콜레스테롤이 높으면 동맥경화를 일으켜 심장마비, 뇌졸중 및 여러가지 심혈관계에 손상을 줌.

위 그림에 콜레스테롤 대사 균형의 개요가 설명되어 있음.
콜레스테롤의 두가지 공급원은 식이 콜레스테롤과 체내에서 합성된 콜레스테롤임. 콜레스테롤은 동물성 음식에 풍부하며 특히 계란의 난황에 높다(난황은 1개 당 250mg의 콜레스테롤을 함유). 섭취한 콜레스테롤의 일부는 흡수되지만, 상당수는 흡수되지 않고대변으로 배설됨.
체내에서 콜레스테롤의 합성에 대해 살펴보면, 대부분의 세포들은 세포막에 필요한 콜레스테롤을 합성하지만 충분하게 합성하지 못하므로 혈액으로부터 공급받음. 체내에서 스테로이드 호르몬을 합성하는 내분비 세포들의 경우도 마찬가지임. 따라서 대부분의 세포들은 혈액으로부터 콜레스테롤을 제거하는 역할을 함. 그러나 간세포와 소화기관내 점막세포는 다량의 콜레스테롤을 합성할 수 있으며, 합성된 콜레스테롤의 대부분은 혈액으로 들어감.
체내 콜레스테롤을 제거하는 간의 기능 중심으로 살펴보는 콜레스테롤 대사균형
첫째, 간은 혈장 콜레스테롤을 받아들여 담즙성분 형태로 소장으로 분비하므로 콜레스테롤을 낮추는 역할을 함. 장간순환으로 되돌아옴.
둘째, 흡수된 콜레스테롤은 간으로 가서 담즙산염으로 합성되어 소장으로 분비됨. 장간순환으로 되돌아옴.
간은 콜레스테롤을 합성하는 기관이며 혈중 콜레스테롤을 제거하기도 하고, 또 담즙 상태로 콜레스테롤을 분비하므로 간은 콜레스테롤의 항상성을 조절하는 주된 기관임. 간은 여러가지 작용에 의해 혈장 콜레스테롤 농도를 정상범위로 유지하지만, 그중 가장 중요한 것은 콜레스테롤 합성작용임. 식사를 통한 콜레스테롤 섭취량이 증가하여 혈장 콜레스테롤 농도가 증가하면, 간에서 콜레스테롤 합성이 저하됨. 그 이유는 혈장 콜레스테롤 농도가 높으면 콜레스테롤 합성에 중요한 HMC-CoA 환원효소의 활성이 저해되기때문.
따라서 콜레스테롤 섭취량이 증가하여 혈장 콜레스테롤 농도가 증가하면 즉시 간에서 합성이 저해되어 혈장 콜레스테롤 농도가 일정하게 유지됨. 반대로 콜레스테롤 섭취량이 저하되면 혈장콜레스테롤 농도가 저하되며, 이로 인해 간에서의 합성이 촉진됨. 그 결과 혈장 콜레스테롤 농도가 원래 수준으로 증가됨. 이때 콜레스테롤 합성이 음성되먹임 조절 방법에 의해 얼마나 민감하게 잘 조절되느냐는 개인적인 차이가 큼. 이처럼 콜레스테롤 합성이 음성되먹임 조절방법에 의해 조절되므로 콜레스테롤 섭취량을 변화시켜도 혈중 콜레스테롤 농도를 낮추기 어려움.
혈중 콜레스테롤 변화
가장 중요한 요인은 식사를 통해 섭취하는 지방산의 양과 형태임. 포화지방(붉은색 고기, 치즈, 전지 우유에 들어있는 동물성 지방)을 많이 섭취하면 혈장 콜레스테롤이 증가하고, 반대로 식물성 지방에 많은 다중 불포화지방산(대두, 옥수수유 등)이나 단일 불포화지방산(땅콩 기름, 올리브유)을 섭취하면 혈장 콜레스테롤이 저하됨. 이들 지방산들은 콜레스테롤의 합성과 배설, 담즙산염으로의 전환에 영향을 줌으로써 혈장 콜레스테롤 농도에 영향을 줌.
현재 콜레스테롤 저하제의 작용. 예를들어 콜레스테롤 합성에 필요한 효소작용을 저해하거나 답즙산염의 장내흡수를 저해함으로서혈장 콜레스테롤을 낮춤.
그러나 혈장 콜레스테롤의 기능이 동일하지 않으며 심혈관계 질병에 미치는 영향도 다양하므로 설명하기가 단순하지 않음. 다른 지방성분들과 마찬가지로 콜레스테롤은 지방단백질의 구성성분으로 혈액을 통해 온몸을 순환함. 지방단백질에는 유미입자, 초저밀도 지방단백질(VLDL), 저밀도 지방단백질(LDL), 고밀도 지방단백질(HDL)이 있음. LDL은 콜레스테롤을 운반하는 주된 지방단백질이며, 콜레스테롤을 온몸의 세포내로 운반함. LDL은 세포막내의 특정 수용체와 결합한 후 세포내섭취에 의해 세포내로 유입됨. LDL과 대조적으로 HDL은 혈액과 조직으로부터 콜레스테롤을 제거하는 역할을 함. 즉 HDL은 콜레스테롤을 간으로 운반하고,그곳에서 담즙을 통해 분비되거나 담즙산으로 전환되며, 스테로이드 호르몬을 합성하는 내분비세포로 콜레스테롤을 운반함.
간과 내분비 세포의 세포막에는 HDL수용체가 많이 들어 있으므로 HDL이 이들 수용체와 결합한 후 세포내로 들어감. LDL콜레스테롤의 농도가 높으면 동맥벽에 콜레스테롤이 많이 축적되고 심장질환 발병율이 증가되기 때문에 LDL콜레스테롤은 해로운 콜레스테롤이라고 하며, 이런 기준으로 볼때 HDL콜레스테롤은 이로운 콜레스테롤이라고 함. LDL콜레스테롤을 해로운 콜레스테롤이라고 하지만, 각종 세포에서 세포막 합성과 교환 또는 부신에서 스테로이드 합성에 필요한 콜레스테롤을 제공하는 중요한 역할을 하므로 적합한 이름인지 아직 애매함.
따라서 동맥경화성 심장질환의 위험을 평가할 수 있는 가장 좋은 지표는 혈장 총콜레스테롤 농도가 아니라 혈장 HDL콜레스테롤에대한 LDL콜레스테롤의 비율임. 이 비율이 낮으면 심장질환의 위험율이 낮음. 심장마비의 위험인자로 알려진 흡연은 혈장 HDL을 낮추는 반면, 과체중인 사람이 체중을 저하시키거나 규칙적으로 운동을 하면 HDL콜레스테롤이 높아짐.
에스트로겐은 LDL을 낮출뿐만 아니라 HDL을 높이므로 폐경전에는 여성의 관상순환게 질환 위험율이 남성보다 낮다. 폐경 후에는 여성의 혈장콜레스테롤과 관상순환계 질환의 발병률이 남성과 유사해짐.
선천적 콜레스테롤 대사이상은 다양한 유형으로 나타남. 예를들면 가족형 고콜레스테롤혈증은 LDL 수용체의 수가 적거나 수용체의 기능이 비정상적일 때 나타나며, 혈액에 LDL이 축적되므로 혈중 LDL농도가 크게 상승함. 그러므로 적절한 치료를 받지 않으면젊은 나이에 동맥경화증이나 심장질환이 나타남.
흡수된 아미노산
흡수된 아미노산의 일부는 간세포로 들어가서 효소와 혈장단백질 등 다양한 종류의 단백질 합성에 이용되거나 탈아미노화 과정에 의해 알카-케토산으로 전환되어 탄수화물합성에 이용됨. 아미노기는 간에서 요소로 전환되어 혈액에 의해 신장으로 운반되어 체외로 배설됨. 알파-케토산은 TCA회로에 의해 분해되어 간세포 활동에 필요한 에너지를 공급하거나, 간세포에서 지방합성에 이용됨.
그러나 섭취한 아미노산의 대부분은 간세포로 들어가지 않고 다른 세로로 들어가서 단백질 합성에 이용됨. 모든 세포에서는 단백질 합성을 위해 아미노산을 계속 필요로 함. 그림 16-1은 단백질 합성과정은 중요하므로 점선 화살표로 나타냄. 이때 식후의 단백질 합성은 공복시 분해된 단백질을 대체하는 정도로만 일어남. 즉 단백질을 과다하게 섭취하여 아미노산의 공급량이 과잉이 될지라도 단백질의 상태로 저장되지 않음. 이점이 포도당이나 지방이 과잉될때 글리코겐이나 지방형태로 저장되는 것과는 뚜렷이 다름.
섭취한 아미노산의 대부분은 체단백질 질량을 유지하기 위해 이용되고, 남은 단백질은 탄수화물이나 지방으로 전환되어 저장됨. 따라서 보통의 성인이 다량의 단백질을 섭취해도 체단백질 질량은 증가되지 않음. 그러나 매일 단백질을 충분히 섭취하면 성장기 어린이나 역도와 같은 근육운동을 꾸준히 하는 성인의 체단백질 질량은 증가될 수 있음.
표 16-1은 식후 흡수상태에서 일어나는 영양소의 대사를 요약함.

공복상태
식후 흡수상태가 지나면 글리코겐, 지방, 단백질의 순합성과정이 끝나고, 이 물질들의 순분해과정이 일어나기 시작함,. 공복시에 일어나는 대사과정은 아래 그림에 요약
공복시에는 소장에서 포도당이 더이상 흡수되지 않지만 포도당을 에너지원으로 이용하는 뇌조직에 포도당을 공급하기 위해 혈당량이 유지되어야 하는 필수적인 문제를 가지고 있는 시기라는 점에서 접근하는 것이 필요함. 이때 혈당량이 정상이하로 저하되면 신경조직활동에 이상이 나타나 정신작용이 저조해지는 경미한 증세가 나타날 수 있으며, 심한 경우 경련, 혼수상태가 되거나 사망함.
혈당량을 유지하는 방법으로 두가지 과정 1) 혈당을 공급해주는 대사반응 2) 세포의 에너지원으로 지방을 이용함으로써 포도당을 절약해주는 대사반응
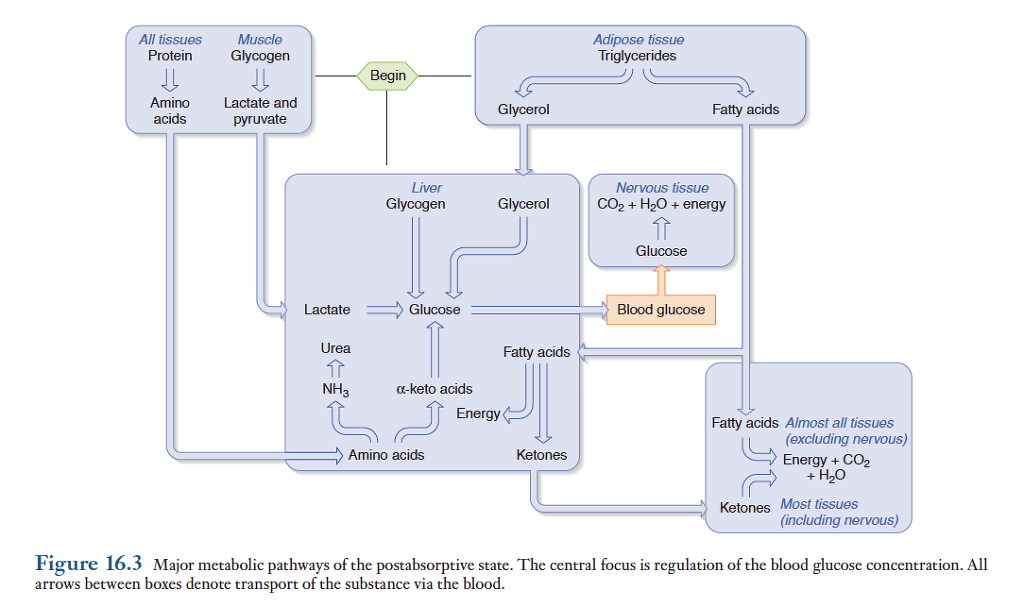
혈당의 공급원
공복시 혈당을 공급해주는 대사과정은 위그림과 같음.
1) 간과 골격근에 저장된 글리코겐이 포도당-6-인산으로 분해되는 과정을 글리코겐분해(glyconeogenesis)라고 함. 간에서 생성된 포도당-6-인산은 효소작용에 의해 포도당으로 분해된 후 혈액으로 들어가 혈당량을 유지시켜 줌. 간 글리코겐은 교감신경계의 흥분과 같이 절적한 자극이 주어졌을때 몇초이내에 분해되기 시작하므로 즉각적으로 혈당량을 유지시켜주는 역할. 그러나 간 글리코겐 저장량은 소량이므로 분해되기 시작한 후 몇시간 이내에 고갈.
골격근에도 간과 유사한 분량의 글리코겐이 저장될 수 있으나, 간과 대조적으로 골격근에는 글리코겐 분해과정의 최종단계인 포도당 6인간을 포도당으로 분해하는 효소가 결여되어 있으므로 근육 글리코겐은 혈당의 공급원으로 작용하지 않음. 골격근내의 포도당 6인산은 해당과정에 의해 ATP, 피루브산 및 젖산으로 분해됨. ATP와 피루브산은 근육세포에서 직접 이용됨. 그러나 젖산은 혈액을 통해 간으로 들어가 혈당량의 유지에 이용됨. 따라서 근육의 글리코겐은 간을 경유하여 간접적으로 혈당량 유지에 이용됨.
2) 지방조직의 중성지방이 글리세롤과 지방산으로 분해되는 것을 지방분해라고 함. 분해된 지방산과 글리세롤은 혈액으로 확산되어 들어감. 글리세롤은 간으로 가서 포도당으로 전환되므로 지방조직의 트리글리세리드로부터 생성된 글리세롤도 공복중 포도당 공급원으로 이용됨.
3) 공복기간 초기에는 단백질이 혈당 공급원으로 작용함. 공복상태가 진행됨에 따라 근육과 다른 조직에서 상당량의 체단백질이 분해되어 혈당량 유지에 필요한 포도당을 공급함. 짧은 기간 동안 정상적으로 일어나는 체단백질 분해는 세포기능을 저하시키지 않으면서도 상당량의 포도당을 공급함. 그러나 공복기간이 연장되어 다량의 체단백질이 지속적으로 분해되면 세포기능이 저하되어 그 기간이 길어지면 질병 또는 사망으로 이어질 수 있음. 단백질이 분해되어 생성된 다량의 아미노산은 혈액으로 들어가 간으로 운반됨. 이들 아미노산은 간에서 알파-케토산으로 분해된 후 포도당 합성에 이용됨.
이와같이 간에서 아미노산과 글리세롤과 같은 전구물질로부터 포도당이 합성되는 과정은 포도당 신생(Gluconeogenesis)라고 알려져 있음. 24시간을 굶을 경우 약 180g의 포도당이 포도당 신생과정에 의해 공급됨. 포도당 신생은 간에서 주로 일어나지만 단식기간이 길어지면 신장에서도 일어남.
포도당 절약작용(지방의 이용)
공복기간동안 간과 신장에서 하루에 약 180g의 포도당이 합성되어 720kcal의 에너지를 공급함. 일반적으로 성인의 하루 에너지 소모량이 1500-3000kcal이므로 포도당 신생만으로 필요한 에너지를 충분히 공급할 수 없음. 따라서 공복상태로 이행되면서 다음과 같은 적응현상이 일어나야 함. 뇌조직을 제외한 대부분의 기관과 조직들은 포도당분해를 최소화하고 지방 이용량을 증가시키고, 지방을 주된 에너지원으로 이용할 수 있도록 대사적으로 적응해야 함. 이러한 대사적 적응현상을 포도당 절약작용(glucose sparing)이라 함.
포도당 절약작용을 가능하게 하는 중요한 대사과정은 지방조직의 트리글리세리드를 글리세롤과 지방산으로 분해하는 지방분해과정(lipolysis)임. 이 과정은 포도당 합성에 필요한 글리세롤을 제공한다는 면에서 중요하지만, 지방산을 유리시켜 신경조직을 제외한 대부분의 조직에 에너지원을 공급한다는 점에서 중요함. 지방산은 혈장알부민과 결합한 상태로 운반되는데 이를 유리지방산(free fatty acid)라고 함. FFA는 신경계를 제외한 여러조직으로 운반되어 다음의 두가지 과정에 의해 에너지를 공급함. 1) 지방산은 베타-산화과정에 의해 전자전달계에서 ATP를 생성할 수 있는 수소원자와 아세틸 조효소를 제공함. 2) 아세틸 조효소는 TCA회로로 들어가 CO2와 H20로 분해됨.
그러나 간은 다른 조직과 달리 특이하게 공복시 지방산으로부터 생성된 아세틸 조효소를 케톤체, 아세톤, 아세토아세트산, 베타-히드록시부티산)로 전환시킴. 케톤체를 아미노산의 대사산물인 알파-케토산과 혼동하지 말것. 공복중에슨ㄴ 케톤체가 혈류를 통해 뇌를 비롯한 여러조직ㅇ로 이동되어 TCA회로에 의해 산화되므로 주요한 에너지 공급원임. 이들 케토체가 혈류를 통해 뇌를 비롯한 여러조직으로 이동되어 TCA회로에 의해 산화되므로 주요한 에너지 공급원임. 이들 케톤체 가운데 아세톤은 휘발성이 크므로 장기간 공복상태인 사람들의 호흡에서 독특한 냄새를 맡을 수 있음.
단식상태에서 지방산과 케톤체가 에너지원으로 이용됨으로써 뇌와 신경계의 에너지원으로 필요한 포도당을 절약해주는 결과가 됨.더욱이 뇌조직은 케톤체를 에너지원으로 사용할 수 있으며, 단식상태가 2-3일 지속될때 케톤체가 혈액에 축적됨에 따라 뇌의 케톤체 이용률이 크게 증가됨. 이러한 변화는 단식상태에 적응하는데 있어 매우 중요한 요소임. 뇌조직이 케톤체를 이용하게 되면 포도달 필요량이 감소되며, 그 결과 포도당 신생을 위해 필요한 아미노산을 공급하기 위한 체단백질 분해를 줄일 수 있기 때문임. 결국더 오랫동안 체 단백질을 보유할 수 있게 되므로 단식기간 중에 나타나는 심각한 조직손상을 피할 수 있음.
표 16-2는 공복상태에서 일어나는 생리적 현상을 요약함.

공복상태에서 글리코겐 분해, 포도당 신생, 에너지원으로 체지방 이용률을 증가시키는 과정이 매우 효율적으로 일어나므로, 며칠간 완전히 단식해도 혈당량은 단지 몇 퍼센트만 저하됨. 한달동안 단식할 경우에도 혈당량은 25%정도 밖에 저하되지 않는다. 그러나 마른 사람의 경우 혈당량 저하가 더 빠름.
2. 식후 흡수상태와 공복상태에서 일어나는 내분비 및 신경조절
식후 및 공복상태에서 일어나는 여러가지 대사과정을 조절하고 통합시키는 내분비 및 신경조절에 대해 살펴보기로 함. 다음의 세가지 질문에 초점을 맞추어 아래 그림에 요약. 1) 식후 흡수상태에서는 단백질, 글리코겐, 트리글리세리드의 순합성은 어떻게 조절되며, 공복상태에서 이 물질들의 순분해는 어떻게 조절되는가? 2) 식후 흡수상태에서 세포가 포도당을 주된 에너지원으로 이용하도록 조절하는 것은 무엇이며, 공복상태에서 포도당 대신 지방을 이용하도록 조절하는 것은 무엇인가? 3) 식후 흡수상태에서 간조직으로 다량의 포도당이 유입되도록 촉진하는 것은 무엇이며, 공복시에 포도당신생과 혈중으로 방출을 촉진하는 것은 무엇인가?
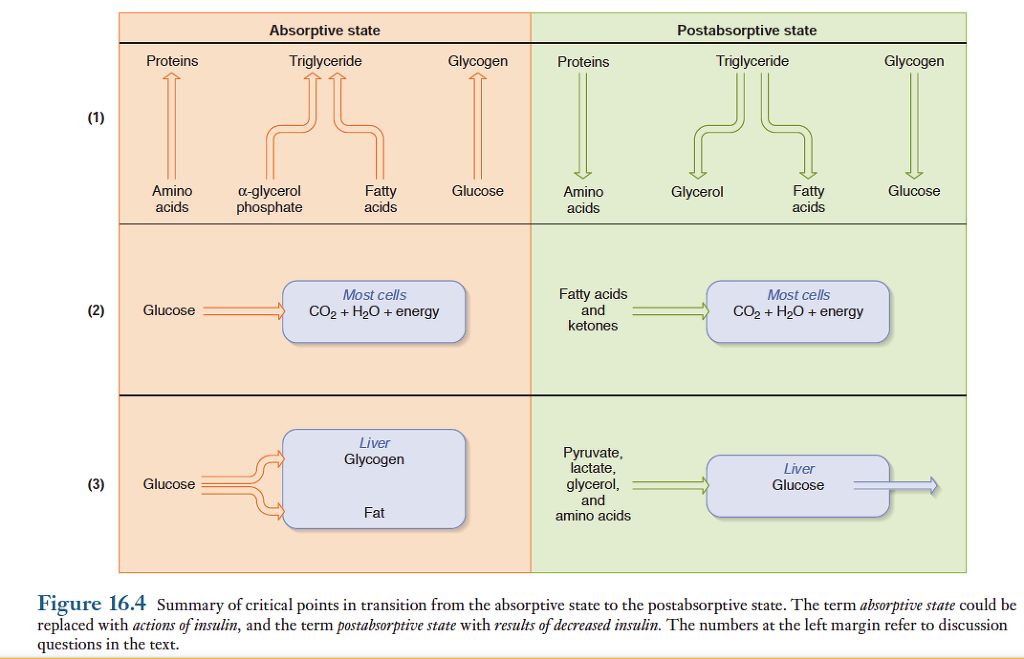
식후와 공복상태에서 일어나는 물질대사의 상호전환을 조절하는 주된 호르몬은 췌장에서 분비하는 인슐린과 글루카곤임. 그 밖에도 부신수질에서 분비되는 에피네프린과 간과 지방조직에서 분포되어 있는 교감신경에 의해 조절됨.
인슐린과 글루카곤은 췌장의 랑게르한스섬에 모여있는 여러가지 다른 종류의 세포가 모여있는데, 인슐린은 베타세포에서, 글루카곤은 알파세포에서 분비됨. 랑게르한스섬에서 분비되는 다른 종류의 분자들의 기능은 아직 밝혀지지 않은 것이 많음. 그 중에서 세타세포에서 분비되는 췌장 소마토스타틴은 시상하부에서 분비되는 소마토스타틴과 동일한 펩티드이며, 뇌하수체 전엽에 작용하여 성장호르몬 분비를 조절함. 인체에서 췌장 소마토스타틴의 생리적 기능은 확실치 않으나, 인슐린과 글루가콘의 분비를 저해함으로써 두가지 호르몬이 과다하게 분비되지 않도록 조절해주는 기능이 있는 것으로 알려짐.
인슐린
인슐린은 물질대사를 조절하는 호르몬 중 가장 중요한 역할을 하며, 때때로 "저장 호르몬"이라고 불림. 혈장내 인슐린 농도는 식후에 증가하고 공복시에 감소함. 인슐린은 근육세포, 지방세포 및 간세포에 주로 작용함. 이들 인슐린의 표적기관에서 인슐린에 의해일어나는 주요 반응을 그림에 요약
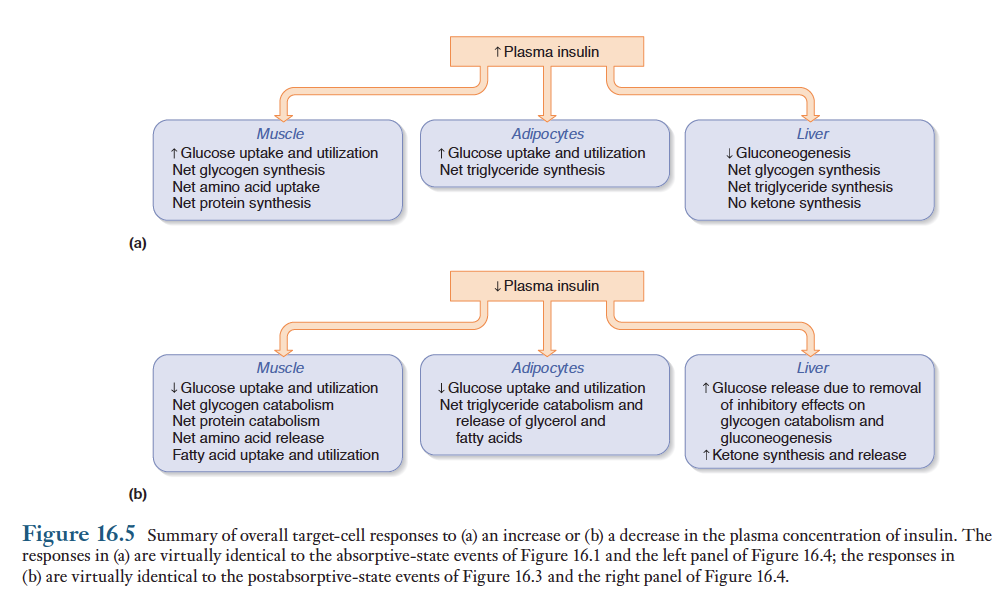
혈장 인슐린 농도의 증가가 식후의 대사과정이 일어나도록 하는 주된 요인으로 작용하며, 혈장 인슐린 농도의 감소는 공복시 대사과정이 일어나도록 작용. 다른 펩티드 호르몬과 같이 인슐린은 표적세포의 세포막에 있는 특수한 수용체와 결합하여 작용함. 인슐린이 수용체에 결합하면 표적세포에 세포막에 있는 운반 단백질과 세포질 내의 효소에 영향을 주는 신호전달과정이 개시됨. 근육세포아 지방세포의 예를들면, 인슐린 농도가 증가되면 세포내에 있는 포도당 운반체인 GLUT-4를 함유하는 소포와 세포막과의 융합이 촉진됨. 그림참조.
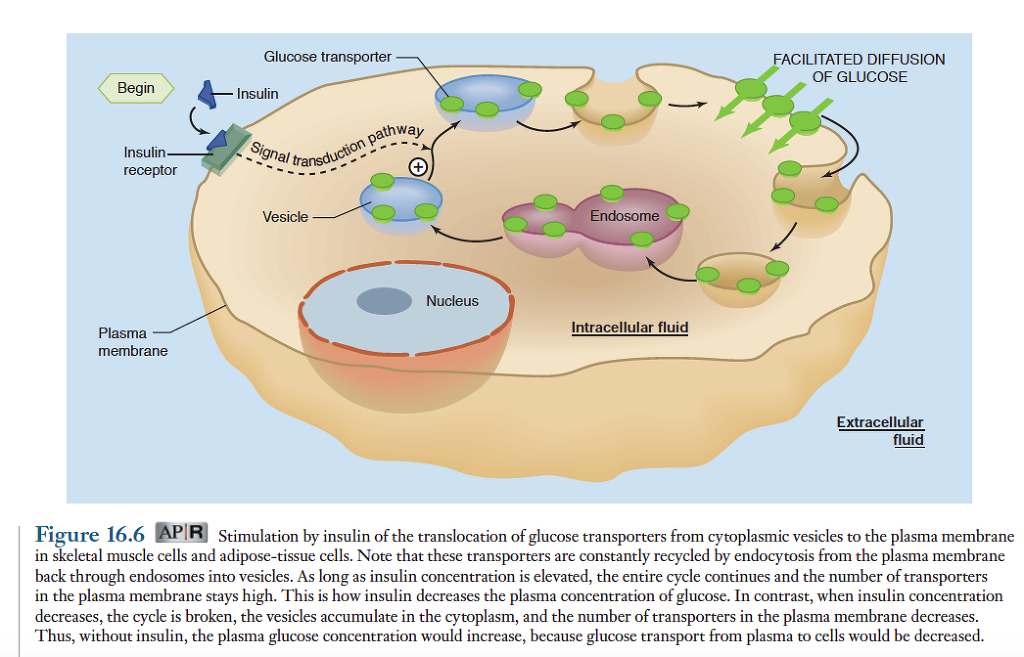
그 결과 세포막에 있는 포도당 운반체의 수는 증가하고, 이로 인해 세포외액의 포도당이 세포내로 촉진확산에 의해 이동되는 속도가 빨라짐. 포도당은 거의 모든 세포에서 촉진확산에 의해 세포내로 들어감. 포도당의 촉진확산에 의한 운반은 여러가지 다양한 형태의 운반체에 의해 이루어지나 근육과 지방조직 세포에 있는 주된 운반체는 GLUT-4로서 이 운반체는 인슐린에 의해 활성이 조절됨. 그러나 뇌에 있는 GLUT 운반체는 이와 다른 종류로서 포도당에 대한 친화력이 매우 높으며 운반체의 활성이 인슐린에 의해 조절되지 않음. 따라서 장기간 단식으로 인해 혈장 인슐린의 농도가 크게 저하되어도 뇌기능에 필요한 포도당을 지속적으로 혈액에서 뇌세포로 운반할 수 있음.
아래 그림에 인슐린에 의한 대사조절을 개괄적으로 제시함.
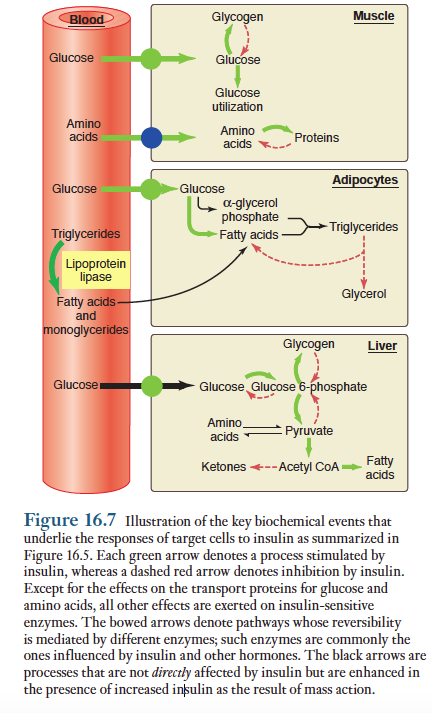
인슐린은 표적세포에서 여러가지 다양한 작용을 함으로써 궁극적인 반응을 일으킴. 예를들면, 근육세포에서 인슐린은 다음과 같은반응을 일으켜서 글리코겐의 합성과 저장을 촉진함. 1) 포도당내의 세포 유입을 증가시킴 2) 글리코겐 합성과정의 속도 조절단계인 글리코겐 합성효소를 활성화함 3) 글리코겐 분해과정의 주요효소인 글리코겐가인산분해효소(glycogenphosphorylase)의 활성을 저해함.
그 결과 인슐린은 근육세포에서 세가지 방법으로 포도당을 글리코겐으로 전환하여 저장되도록 촉진함. 이와 유사한 인슐린은 다음과 같은 방법으로 근육세포에서 단백질 합성을 촉진함. 1) 근세포막의 아미노산 운반체 수를 증가시켜 아미노산의 유입량을 증가시킴. 2) 단백질 합성에 필요한 리보솜에 있는 효소들을 활성화 함. 3) 단백질 분해효소들의 작용을 저해함.
인슐린 분비의 조절
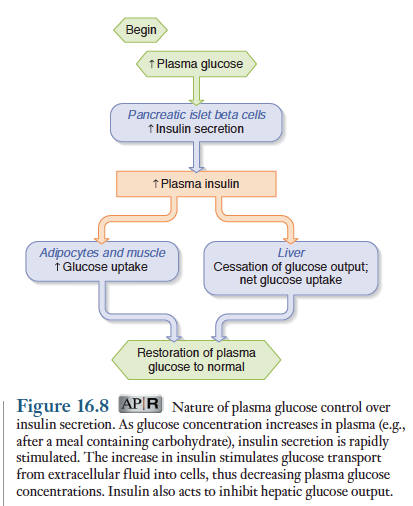
인슐린 분비를 조절하는 주된 요인은 혈당량
식후 혈당량이 상승하면 랑게르한스섬 베타세포를 자극하여 인슐린 분비, 혈당량이 저하되면 인슐린 분비자극이 제거됨. 아래 그림 되먹이기 기전
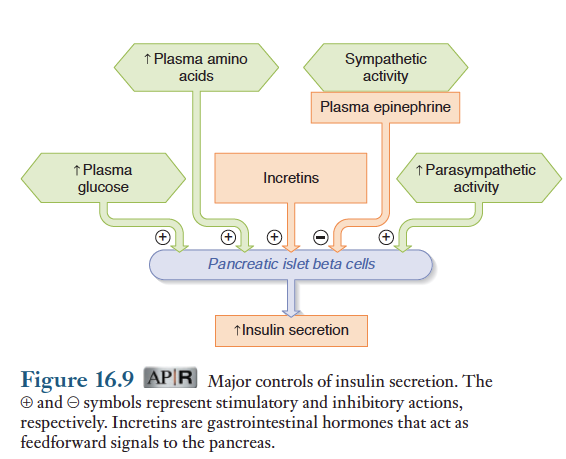
혈당량 이외에 인슐린 분비자극하는 요소
예를들면 혈중 아미노산 농도 증가, 소화기관에서 분비되는 인크레틴이라는 호르몬은 포도당에 대한 인슐린 반응을 증폭시키는 역할. 인크레틴 가운데 글루카곤-유사 펩티드 1(GLP-1)은 가장 주요 호르몬임. 인크레틴은 식후에 인슐린 분비를 촉진함으로써 인슐린 분비가 혈당량에 의해서만 촉진되는 것보다 더많이 분비되도록 조절하는 것이 가능함. 그 결과 식후에도 혈당량이 지나치게 증가하지 않게 되므로 신장의 네프론에서 완전히 재흡수 될 수 잇는 수준으로 혈당량이 유지됨. 식후 흡수상태가 지난 후 혈당량이 공복상태보다 높은 상태에서 인크레틴 분비가 저하됨으로써 인슐린 분비로 인해 올 수 있는 저혈당을 방지함.
마지막으로 랑게르한스섬에 분포되어 있는 자율신경도 인슐린 분비에 영향을 줌. 식사하는 동안 부교감신경이 흥분되면 인슐린 분비가 촉진되는데, 이 조절방법도 또 다른 식후 촉진조절에 속함. 이와 대조적으로 랑게르한스섬에 분포되어 있는 교감신경이 흥분하거나 부신수질에서 에피네프린 분비가 증가되면 인슐린 분비를 저해함. 저혈당이나 스트레스, 운동에 대한 신체 반응 즉 교감신경이 흥분되는 상황에서 조절과 관련된 내용은 이 장의 뒷부분에서 설명.
글루카곤
글루카곤은 췌장의 알파세포에서 분비되는 펩티드 호르몬. 글루카곤은 간조직에 주로 작용하며, 인슐린과 길항작용을 함. 아래 그림.
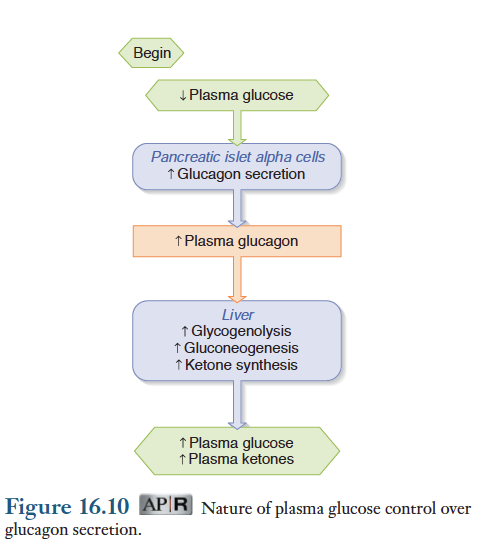
간조직에서 글루카곤은 1) 글리코겐을 분해 2) 포도당 신생작용을 증가 3) 케톤체 합성을 증가시킴. 따라서 글루카곤이 분비되면 혈중에 포도당과 케톤체 농도가 증가되므로 공복시에 중요하며 저혈당을 예방함. 그러나 지방세포에 대한 글루카곤의 작용은 분명치 않음.
글루카곤 분비를 자극하는 요인은 저혈당 상태임. 즉 혈당량이 저하되면 글루카곤이 분비되어 글리코겐 분해와 포도당 신생작용을촉진하여 혈당량을 정상적으로 유지시켜줌. 동시에 글루카곤은 뇌조직이 사용할 수 있는 케톤체를 공급함.
교감신경은 랑게르한스섬을 자극하여 글루카곤 분비를 촉진하는 반면에 인슐린의 분비를 억제함.
간조직과 지방조직에 대한 에피네프린과 교감신경의 조절작용
에피네프린과 교감신경은 췌장 랑게르한스섬에서의 인슐린의 분비를 억제하고 글루카곤 분비를 촉질함. 또한 에피네프린은 영양소대사에도 직접적으로 영향. 1) 간과 근육에서 글리코겐 분해 촉진 2) 간의 포도당 신생과정 촉진 3) 지방세포의 지방분해 촉진.
교감신경이 흥분했을때 간과 지방세포에 나타나는 영향은 혈중 에피네프린 농도가 증가하였을때와 동일함.
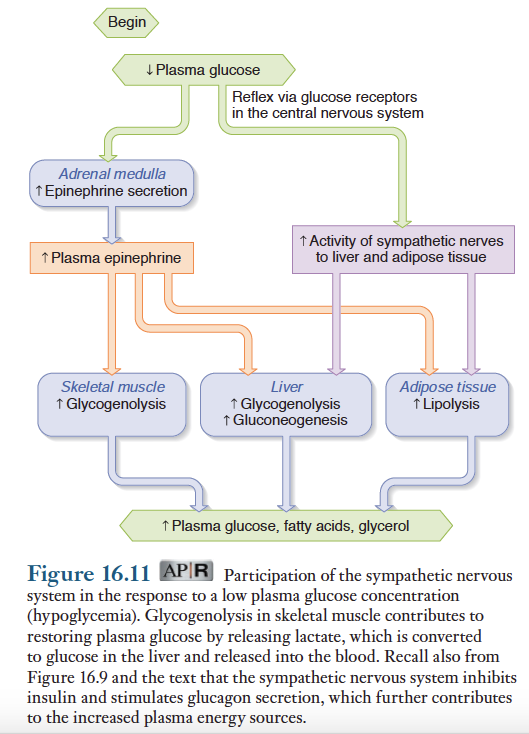
지방조직에서 에피네프린은 호르몬-민감성 리파아제(hormone-sensitive lipase, HSL)를 활성화 함. 일단 HSL이 활성화되면 트리글리세리드를 유리지방산과 글리세롤로 분해함. 이 두 물질은 혈중으로 유리되며, 지방산은 에너지원으로 직접 이용되고 글리세롤은포도당 신생에 이용됨. 식후에 분비되는 인슐린은 HSL의 활성을 저해. 따라서 교감신경이 흥분하면 인슐린의 작용과 반대되는 대사과정이 활성화됨. 즉 혈중 포도당, 글리세롤 및 지방산의 농도가 증가함.
교감신경의 작용으로부터 예상되듯이, 저혈당 상태는 간과 지방세포에 대한 에피네프린과 교감신경 작용이 증진. 교감신경과 에피네프린이 글루카곤 분비를 촉진한다는 점에서 동일하지만, 이에 관여하는 수용체나 경로는 완전히 다름. 혈중 포도당 농도가 저하되면 중추신경계(및 간조직)에 있는 포도당-민감성 세포들이 반사적으로 부신수질, 간 및 지방조직의 교감신경을 자극함. 그 결과 나타나는 대사적 적응현상은 저혈당 상태에서 글구카곤이 분비되었을때 나타나는 반응과 동일함. 즉 혈당량이 정상수준으로 증가되면, 세포의 에너지원으로 이용될 수 있는 지방산이 공급됨.
코르티솔
코르티솔은 부신수질에서 분비되는 글루코코르티코이드로서 단식상태에 필요한 적응반응에 대해 "허용작용"을 함. 단식상태에서는포도당신생과 지방분해가 촉진. 그러나 코르티솔이 정상적으로 분비되지 않을 경우, 이러한 대사 전환이 정상적으로 이루어지지 않음. 다시말하면 단식상태에서 혈중 코르티솔 분비가 크게 증가될 필요는 없지만, 혈중 코르티솔 농도가 적절해야만 포도당 신생과 지방분해에 필요한 효소들(예, HSL)의 농도가 유지됨.
따라서 코르티솔이 전혀 분비되지 않는 사람이 단식을 하게 되면 심한 저혈당을 초래하여 뇌기능에 심각한 손상을 가져올 수 있음.또한 혈중 코르티솔 농도가 상승하면 스트레스를 받는 상황과 마찬가지로 코르티솔은 허용 작용 이상의 역할을 함. 코르티솔 분비가 높을때 나타나는 대사적 변화는 공복상태에서 나타나는 대사적 변화와 유사함. 아래 표 설명.

사실상 코르티솔의 역할은 근육과 지방세포의 인슐린에 대한 민감도를 저하시켜서 공복시에 혈당이 유지돕도록 도우며, 그 결과 뇌가 에너지원을 적절하게 공급받도록 돕는 것임. 이와같이 코르티솔은 인슐린에 대한 길항 작용을 나타내는 글루카곤이나 에피네프린과 비슷한 작용을 하는 또 다른 호르몬이라 할 수 있음. 실제로 유전적인 원인이나 임상적인 문제로 인해 혈중 코르티솔 농도가 비정상적으로 매우 높은 사람들은 2형 당뇨환자와 같이 인슐린에 대해 부적절하게 반응하는 사람들과 유사한 증세를 나타냄.
성장 호르몬
성장호르몬의 주된 생리적 기능은 인슐린-유사성장인자 1(insulin-like growth factor-1)을 매개로 하여 성장과 단백질 합성을 촉진하는 작용임. 이 기능들에 비해 탄수화물이나 지방대사에 대한 성장호르몬 작용은 사소한 편임. 그러나 코르티솔과 마찬가지로 성장 호르몬이 심하게 부족하거나 지나치게 많이 분비되면 지방과 탄수화물 대사가 비정상적으로 일어남. 성장 호르몬이 지방과 탄수화물의 대사에 미치는 영향은 코르티솔과 유사하고 인슐린과 반대영향을 가져오므로 단백질 대사에 미치는 영향과는 대조적임.
성장 호르몬은 1) 지방세포가 지방분해를 일으키는 자극에 대해 더 잘 반응하게 해주며 2) 간에서 포도당신생작용을 증가시키고 3) 간과 근육에 포도당 유입을 증가시키는 인슐린의 작용을 억제함. 이 세가지 작용을 성장 호르몬 과다분비에 의해 말단비대증인 사람은 제 2형 당뇨환자와 같이 인슐린 내성이 있은 사람과 유사한 증세를 나타냄.
호르몬에 의한 대사조절 요약
인슐린은 풍요의 호르몬이라 할 수 있음. 인슐린 분비가 식후에 증가되고, 공복시 감소됨으로써 대사과정을 적절하게 변화시킴. 글루카곤, 에피네프린, 간과 지방조직에 분포되어 있는 교감신경, 코르티솔 및 성장 호르몬은 혈당조절에 있어서 인슐린과 반대 작용을 함. 아래 표 참조.
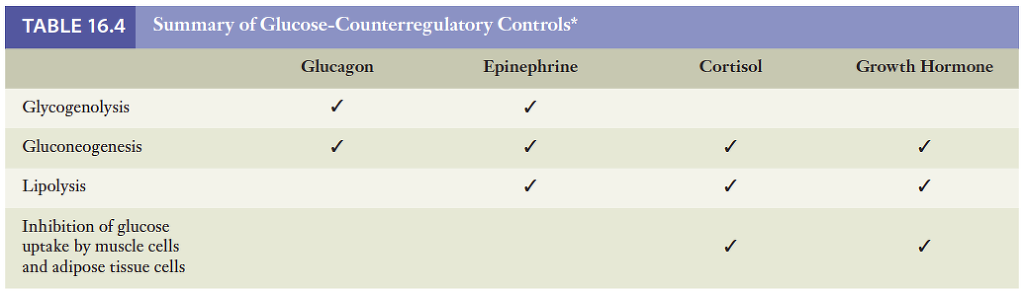
글루카곤과 교감신경은 저혈당 방지에 가장 중요한 역할을 함. 식후흡수 상태에서 공복상태로 바뀌어도 코르티솔과 성장 호르몬의분비량은 이에 맞추어 변화되지 않음. 그러나 공복상태에의 지방대사와 탄수화물 대사가 정상적으로 이루어지기 위해서는 이 두 호르몬의 혈중 농도가 일정수준으로 유지되어야 하며, 이 두 호르몬 중 어느 하나라도 과다하게 분비되면 혈당량이 비정상적으로 높아짐. 수면 중에 성장 호르몬 분비가 증가되며 잠에서 깨기 직전에 코르티솔 분비가 증가됨으로써 수면중의 혈당을 유지시켜줌.
3. 운동과 스트레스 상태에서의 에너지 항상성 유지
운동중에는 근육수축에 필요한 에너지를 공급하기 위해 다량의 에너지원이 소모되어야 함. 이들 에너지원으로는 혈장 포도당과 지방상 및 근육내의 글리코겐이 이용됨. 운동 중에 이용되는 혈장 포도당은 간에서 글리코겐 분해와 포도당 신생에 의해 공급됨. 또한 운동시 지방조직에서 다량의 지방산이 분해되며 그 결과 글리세롤과 지방산이 혈중으로 방출됨. 이때 글리세롤은 간으로 가서 포도당 신생에 이용되며, 지방산은 운동중인 근육에 필요한 에너지원으로 이용됨.
운동중에 혈당량은 어떤 변화가 일어나는가?
단시간에 걸친 운동이나 강도가 약하거나 보통 정도인 운동을 할때에는 혈당량은 거의 변화 없으며, 높은 강도의 운동을 단시간 시행할때 혈당량은 약간 높아지기도 함. 그러나 운동시간이 길어지면(90분이상) 혈당량이 감소되기는 하지만 그 감소량은 25%이하임.
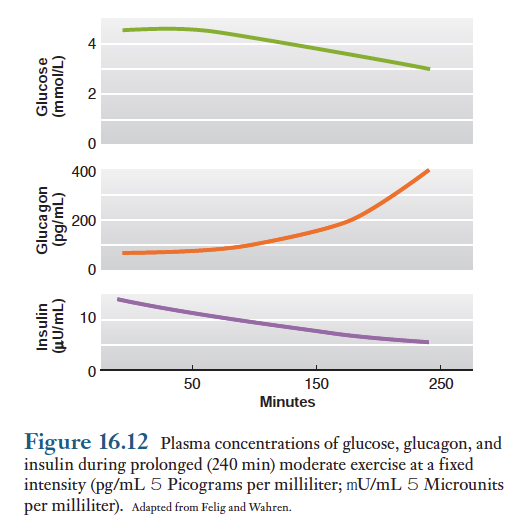
운동시 간에서 혈중으로 방출되는 포도당의 양은 운동에 필요한 포도당에 비례적으로 증가함. 이런 변화는 운동초기에는 약간의 잠복기를 거쳐 시작됨. 운동 중인 사람의 대사상태는 공복상태와 유사하며(간에서 포도당 신생증가, 트리글리세리드분해 증가, 지방산 이용증가), 운동중의 호르몬에 의한 대사 조절도 공복상태인 사람과 동일함. 운동중에는 인슐린 분비가 감소되고 글루카곤 분비가 증가되며, 이 두 호르몬의 혈중 농도가 대사를 조절하는 주된 역할을 함. 또한 교감신경계의 활성이 증가되고 에피네프린, 코르티솔 및 성장호르몬의 분비가 증가함.
그렇다면 운동할때 글루카곤의 분비를 촉진하고 인슐린 분비를 감소시키는 요인은 무엇인가?
한가지 요인으로는 혈당량의 저하를 들 수 있는데, 운동중에는(장시간 운동) 혈당량이 약간 저하되며 이로 인해 호르몬 분비에 영향을 줌. 또한 운동의 강도와 관계없이 모든 운동이 혈중 에피네프린 농도를 상승시키고, 췌장에 분포되어 있는 교감신경의 활동을증가시킴. 이와같이 교감신경이 흥분하면 직접적으로 간과 지방조직에 작용하여 에너지 원으로 이용되는 영양소를 동원하고, 간접적으로는 인슐린의 분비를 억제하고 글루카곤의 분비를 촉진하는 작용을 함. 이와같이 교감신경을 조절하는 것은 혈당의 변화가 아니라 운동에 의한 일종의 신경반응으로서 중추신경계에 의해 조절됨.
운동중에 일어나는 반응 가운데 공복 중에 일어나는 반응과 완전히 다른 점은 근육에서의 포도당 유입량과 이용량이 운동중에는 증가되지만 공복중에는 현저히 감소된다는 점임. 이처럼 운동중에는 혈장 인슐린 농도가 낮아지며 근육세포로 포도당 유입을 감소시키는 코르티솔과 성장호르몬 농도가 높음에도 불구하고 다량의 포도당이 근세포막을 통해 촉진확산으로 이동될 수 있는 것이 어떻게 가능한가? 지금까지 잘 알려지지 않은 방식인 근수축 작용에 의해 세포질에 있는 포도당 운반체가 세포막으로 이동되어 세포막을 통한 포도당 운반을 증가시킴. 따라서 운동중인 근육은 휴식중인 근육보다 더 많은 포도당을 더 적은 양의 인슐린 분비로 근육세포내로 운반할수 있음.
운동상태와 공복상태 이외에도 인슐린 분비가 감소되고 글루카곤 분비, 교감신경 활성 및 코르티솔과 성장 호르몬분비가 증가되는경우가 있음. 즉 신체적 및 감정적으로 다양한 스트레스를 받는 상황에서 이와 유사한 호르몬 분비양상이 나타나므로 위협적인 스트레스를 대비하여 대사적인 준비를 할 수 있게 됨. 또한 인슐린 분비가 저하되고 코르티솔 분비가 증가되면 체단백질이 아미노산으로 분해되어 포도당 신생에 이용되므로 필요한 에너지를 공급해줄 뿐만 아니라 상해를 입을 경우 조직회복에 필요한 아미노산을공급해줌.
강도가 높은 매일 오랫동안 운동을 하면 인체에 스트레스를 줌. 이 경우 신체에 비필수 기능들이 중단됨으로써 더 많은 영양소들이근육으로 가서 우선적으로 이용될 수 있게 됨. 비필수 기능의 한 예는 생식기능임. 따라서 격렬한 연습을 통해 운동에 단련중인 청소년들의 경우 사춘기가 지연됨. 이와 유사하게 격렬하게 운동연습을 하는 여성들은 일시적으로 불임상태(운동-유발성 무월경)가 되기도 함. 발레 선수와 같이 체중감소와 격렬한 연습이 요구되는 직업에서 이런 상태를 볼 수 있음. 남성들도 운동을 심하게 하면생식능력이 저하되는지는 확실하지 않으나 생식능력에 이상이 없다는 증거들이 많이 있음.