활동전위
활동전위, action potential, AP, 동작전위
근세포·신경세포 등 일부 세포의 막 전위가 일시적으로 크게 상승하는 현상을 가리킨다. 세포막에는 각종 이온 통로와 수송 단백질이 있어 나트륨·칼륨 등 이온의 농도가 안팎이 서로 다르다. 이온 조성의 차이로 말미암아 안정 상태에서 세포막 안쪽의 전위는 바깥쪽에 비해 보통 -60∼90 mV 정도 낮은데, 이 전위차를 휴지 전위라고 한다. 일부 흥분성 세포(영어: excitable cell)는 자극을 받거나 스스로 흥분하면 막 전위를 빠르게 역전시켜 순간적으로 휴지 전위보다 훨씬 높은 +30~40 mV에 이르도록 할 수 있다. 이것이 활동전위이다. 활동전위에 의한 막 전위 변동은 수 밀리초(ms) 정도만에 원래대로 회복되므로 스파이크 전위(spike potential)라고도 하며, 회복기에 보이는 느린 변화인 후전위(after potential)와 구별된다.
활동전위는 막 전위 값이 원래대로 돌아오면서 그대로 사라지는 것이 아니라, 주변으로 전파되어 새로운 활동전위가 만들어지도록 한다. 이에 따라 국소전류가 발생하여 1∼100 m/s의 속도로 흥분이 전달된다. 이를 율리우스 베른슈타인(Julius Bernstein)이 탈분극 현상으로 설명하였다. 이처럼 세포막 전체로 확산·전파되므로 활동전위는 세포 내 신호 전달에 활용될 수 있다.
활동전위는 신경세포와 일부 신경내분비세포, 근세포에서 전기적 신호를 전달하는 방법으로 사용된다. 끈끈이주걱·활수초 등 일부 식물 세포에서도 활동전위가 관찰된다.
활동전위는 크게 전압 개폐 나트륨 통로, 전압 개폐 칼륨 통로 및 다른 이온 채널에 의해 일어난다.
대부분의 세포에서 전압은 일정한 편이지만 전기적으로 활동적인 세포는 전압이 변화한다. 전압은 특정한 경우에 매우 빠르게 오르락내리락하고, 그 형태는 정형화된 양식을 따른다. 이 변화 주기를 활동전위라 한다. 활동전위가 지속되는 시간은 넓은 범위에 걸쳐 변화하여 결과적으로 아날로그 신호에 속한다. 동물의 뇌세포에서 변화는 ms 단위로 일어난다. 기타 세포에서 변화 주기는 수 초에 걸쳐 일어날 수 있다.
동물세포의 전기적 특성은 막 구조에 의해 결정된다. 세포막은 단백질 분자가 박혀있는 지질층으로 이루어져 있다. 지질층은 전하를 띤 이온에 저항이 커서 부도체 기능을 한다. 반면에 막 단백질은 채널을 형성하여 이온이 막을 통과할 수 있게 한다. 몇몇 분자는 특정한 이온을 막 안팎으로 능동수송할 수 있다.
뉴런에서의 과정
동물의 신체 조직을 이루는 세포는 세포막 안팎으로 막 전위를 유지한다. 전기적 극성을 유지하는 것은 막에 박혀있는 이온 펌프와 이온 채널이다. 뉴런의 부분별로 막에 있는 이온 채널이 달라 가지돌기, 축삭, 세포체가 각각 전기적 특성이 다르다. 뉴런 어떤 부분의 막은 흥분성(활동전위를 생성할 수 있음)이고 어떤 부분은 그렇지 않다. 축삭 언덕(시작 마디, 축삭과 세포체가 만나는 부분) 뒤부터 흥분성 부위라는 연구 결과가 있으나, 대부분의 사례에서는 세포체와 축삭 또한 흥분성이다.
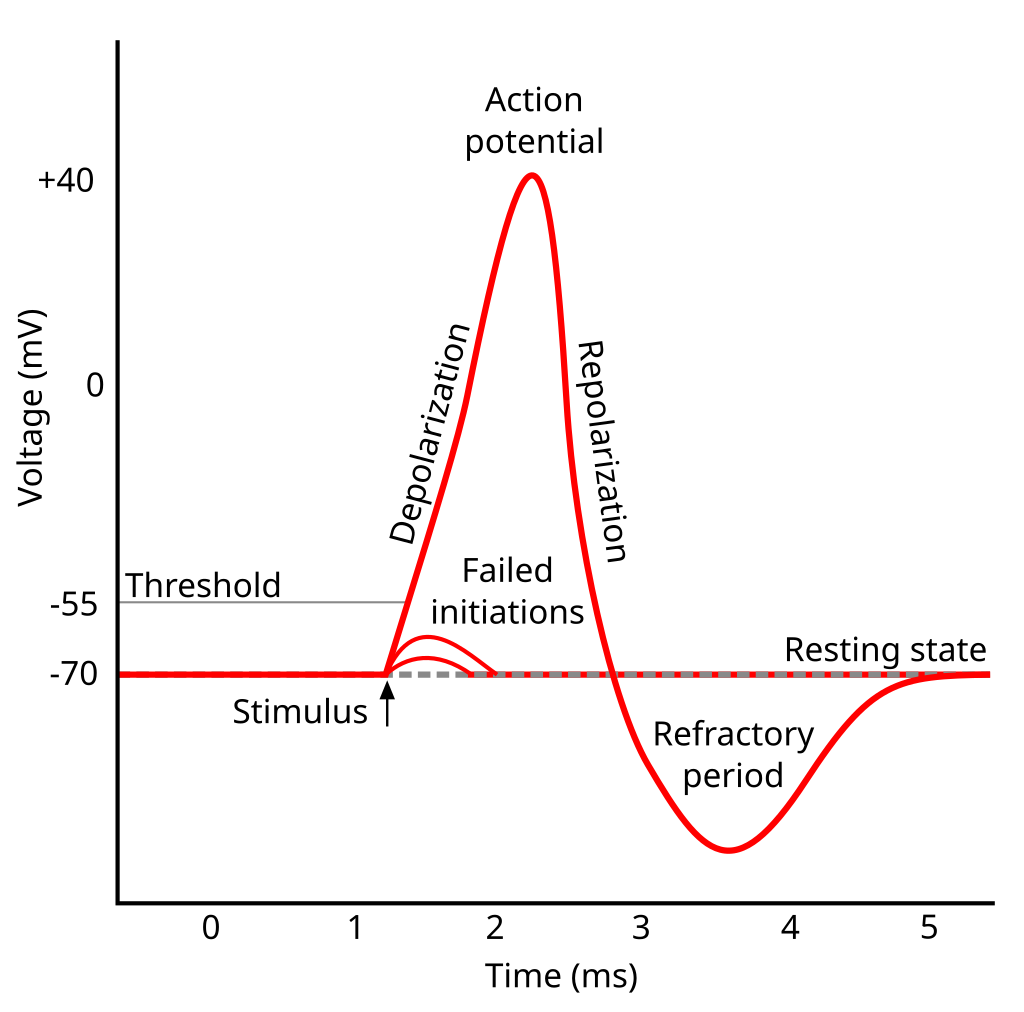
흥분성 막 부분에서 중요한 막 전위는 세포에 아무런 동요가 없을 때 막 전위인 휴지 전위와 그보다 높은 값인 역치 전위이다. 전형적인 뉴런 축삭 언덕에서 휴지 전위는 -70 mV, 역치 전위는 -55 mV 내외이다. 뉴런에 시냅스 자극이 오면 막은 탈분극(막 전위가 상승)이나 과분극(감소)한다. 활동전위는 막이 충분히 탈분극하여 역치까지 도달하면 일어난다. 역치를 넘어 탈분극이 일어나면 막 전위는 급격히 상승(+100 mV까지 도달하기도 한다)하고, 다시 급격히 감소하여 휴지 전위 아래까지 떨어졌다가 일정 시간 유지한 후 휴지 전위로 돌아온다. 활동전위의 형태는 일정하게 고정되어 있다. 즉 어떤 세포에서 활동전위는 같은 크기(amplitude)와 시간 주기로 일어난다(예외적인 경우를 이후에 다룸). 대부분의 뉴런에서 활동전위 전체 과정은 천 분의 일 초 내에 일어난다. 여러 뉴런이 초당 최대 10-100회의 속도로 활동전위를 일정하게 내보낸다. 그렇지 않은 뉴런은 수 분 이상 아무런 활동전위를 발생하지 않기도 한다.
생물물리학적 기초
축삭을 따라 전파되는 활동전위
활동전위는 세포막에 있는 특별한 전압 개폐 이온 채널에 의해 일어난다. 전압 개폐 이온 채널은 막에 박혀있는 단백질 집단으로 세 가지 특성을 지닌다.
한 개 이상의 입체형태를 가진다.
최소 한 개의 입체형태는 특정한 이온이 막을 통과하도록 한다.
한 입체형태에서 다른 입체형태로 전이(transition)는 막 전위의 영향을 받는다.
전압 개폐 이온 채널은 특정한 막 전위값에서 열리고 닫힌다. 대부분의 경우 막 전위와 채널 상태 사이의 관계는 확률론적이고 개폐에 시간차가 존재한다. 이온 채널은 입체형태를 예측할 수 없는 시각에 바꾼다. 막 전위는 전이 속도와 단위 시간당 각 입체형태일 확률을 결정한다.
전압 개폐 이온 채널은 양성 피드백 고리로 활동전위를 만들어낸다. 막 전위는 이온 채널의 상태를 통제하고 이온 채널은 막 전위를 통제한다. 따라서 특정 상황에서 막 전위가 올라가면 이온 채널이 열리고, 이로 인하여 막 전위가 더욱 상승한다. 활동전위는 양성 피드백 주기가 폭발적으로 진행될 때 일어난다. 활동전위의 시간과 크기 궤도는 활동전위를 생산하는 전압 개폐 이온 채널의 생물물리학적 특성이 결정한다. 다양한 채널이 활동 전위를 만들어내는 양성 피드백 기능을 한다. 전압 개폐 나트륨 채널은 신경 전도와 관련된 빠른 활동전위를 만든다. 근육 세포와 몇몇 뉴런에서 일어나는 느린 활동전위는 전압 개폐 칼슘 채널이 담당한다. 각각의 채널에는 전압 민감도와 시간에 따른 움직임이 다른 다수의 변형체(variant)가 존재한다.
가장 널리 연구되는 전압 의존성 이온 채널은 빠른 신경 전도에 관련된 나트륨 채널이다. 나트륨 채널과 활동전위의 생물물리학적 연구로 노벨상을 수상한 앨런 로이드 호지킨과 앤드루 헉슬리의 이름을 따 나트륨 채널을 호지킨-헉슬리 나트륨 채널이라고도 부르고, 더욱 단순하게는 NaV 채널(V는 전압을 의미)이라고 부른다. NaV 채널은 불활성형(닫힌형, deactivated state), 활성형(열린형, activated state), 차단형(inactivated state) 등 세 가지 상태로 존재할 수 있다. 채널은 활성형 상태일 때 나트륨 이온에 대해서만 투과성이 있다. 막 전위가 낮을 때 채널은 대부분의 시간 동안 불활성형으로 존재한다. 막 전위가 특정 수준 이상으로 올라가면 채널이 활성형으로 전이될 확률이 증가하고, 막 전위가 높을수록 확률은 더 커진다. 채널이 활성화되고 나면 차단형으로 변화하고, 그 상태로 일정 시간 유지된다. 막 전위가 다시 낮아지면 채널은 불활성형으로 돌아간다. 결론적으로 활동전위 동안 채널이 거치는 주기는 불활성형 → 활성형 → 차단형 → 불활성형 순이다. 이 순서는 채널 전체를 기준으로 평균적인 행동이고, 이론적으로 각각의 채널은 한 시점에서 전이 상태가 모두 다를 수 있다. 다만 채널이 차단형에서 곧바로 활성형으로 바뀌는 것은 거의 일어나지 않는다. 차단형에 있는 채널은 불활성형으로 되돌아갈 때까지 불응기(무반응성, refractory)를 거친다.
이러한 NaV의 동역학적 특성의 결과 NaV는 천이 매트릭스(transition matrix)의 지배를 받는다. 채널 자체가 전압을 결정하는 중요 역할을 하기 때문에, 채널이 존재하는 계의 전체적인 동역학을 계산하기는 상당히 어렵다. 호지킨과 헉슬리는 이온 채널의 상태를 나타내는 매개변수(parameter)에 대한 미분 방정식, 호지킨-헉슬리 방정식을 세웠다. 호지킨-헉슬리 방정식은 계속 변형되었지만 활동전위를 연구하는 생물물리학의 이론적인 출발점이 되고 있다.
막 전위가 올라감에 따라 나트륨 이온 채널이 열려 세포 내로 나트륨 이온이 들어온다. 칼륨 이온 채널이 그 뒤를 이어 열려 칼륨 이온이 세포 바깥으로 빠져나간다. 나트륨 이온의 유입은 양전하를 띤 양이온의 농도를 증가시켜 휴지 전위보다 높은 전위 상태인 탈분극을 유발한다. 활동전위가 최고에 달했을 때 나트륨 채널은 닫히고 칼륨 채널은 계속해서 열려있어 칼륨이 세포 밖으로 나간다. 칼륨 이온의 유출로 막 전위가 감소하고 세포는 과분극된다. 이 지점에서는 칼륨 흐름이 나트륨 보다 많아 전위는 약간 상승하고 정상적인 휴지 전위(대개 -70 mV)에 도달한다. 휴지 전위보다 15 mV 높은 역치 전위를 지나 더 상승하면 나트륨 흐름이 더 우세하다. 그 결과 양성 피드백으로 나트륨 흐름은 나트륨 채널을 더 많이 활성화한다. 따라서 세포는 발화하여 활동 전위를 만든다.[note 1] 세포의 활동전위가 발생하는 빈도를 발화율이라 한다.
전압 개폐 채널을 열어서 생기는 이온의 흐름은 대개 활동전위를 만드는 초기 흐름보다 크다. 따라서 활동전위의 크기와 지속 시간, 형태는 자극의 크기나 지속 시간이 아니라 흥분하는 막의 특성에 대부분 의존한다. 이러한 실무율 특성은 수용기 전위, 전기긴장성 전위, 시냅스 전위 등 자극의 세기에 따라 증감하는 다단계 전위와 구별된다. 활동전위는 전위 개폐 채널과 유출 채널(leak channel)의 종류, 채널 분포, 이온 농도, 막 전기 용량, 온도 및 기타 요소에 따라 다양하다.
활동전위에 관여하는 주요 이온은 나트륨과 칼륨 양이온이다. 나트륨 이온은 세포 안으로, 칼륨 이온은 세포 밖으로 이동하여 평형을 유지한다. 상대적으로 적은 이온 이동만으로도 막 전위를 극적으로 변화시킬 수 있다. 따라서 활동전위 동안 이온 교환으로 변하는 막 안팎의 이온 농도는 무시할 수 있다. 막을 건너는 이온 중 소수는 다른 이온 수송체와 함께 정상 이온 농도비를 유지하는 나트륨-칼륨 펌프에 의하여 다시 되돌아간다. 칼슘 양이온과 염소 음이온은 심장 활동전위와 단세포 조류 Acetabularia의 활동전위 등 소수의 활동전위 유형과 연관이 있다.
활동전위는 막의 흥분성 부분에서 국소적으로 만들어지지만, 휴지 상태의 이온 흐름도 인접한 막의 연장선에서 활동전위를 유발하고 도미노처럼 퍼지게 할 수 있다. 수동적으로 퍼지는 전기긴장성 전위와 반대로, 활동전위는 흥분이 막을 따라 뻗어가면서 계속 새로 만들어지고, 감소하지 않는다. 축삭에서 수초가 있는 부분은 흥분성이 아니고 활동전위를 만들지 않아 신호가 전기긴장성 전위처럼 수동적으로 전파된다. 일정한 간격으로 수초가 없는 부분, 즉 랑비에결절은 활동전위를 만들고 신호를 빠르게 도약 전도한다. 도약 전도는 축삭 지름이 작아도 신호의 속도를 높여주는 효과가 있다. 축삭 말단이 탈분극되면 신경전달물질이 시냅스 틈으로 방출된다. 신피질 어디에나 존재하는 피라미드 뉴런의 가지돌기에서는 활동전위가 거꾸로 전파되기도 하는데, 이 현상은 돌기-시기 의존성 가소성에 영향을 미치는 것으로 추정된다.
활동전위
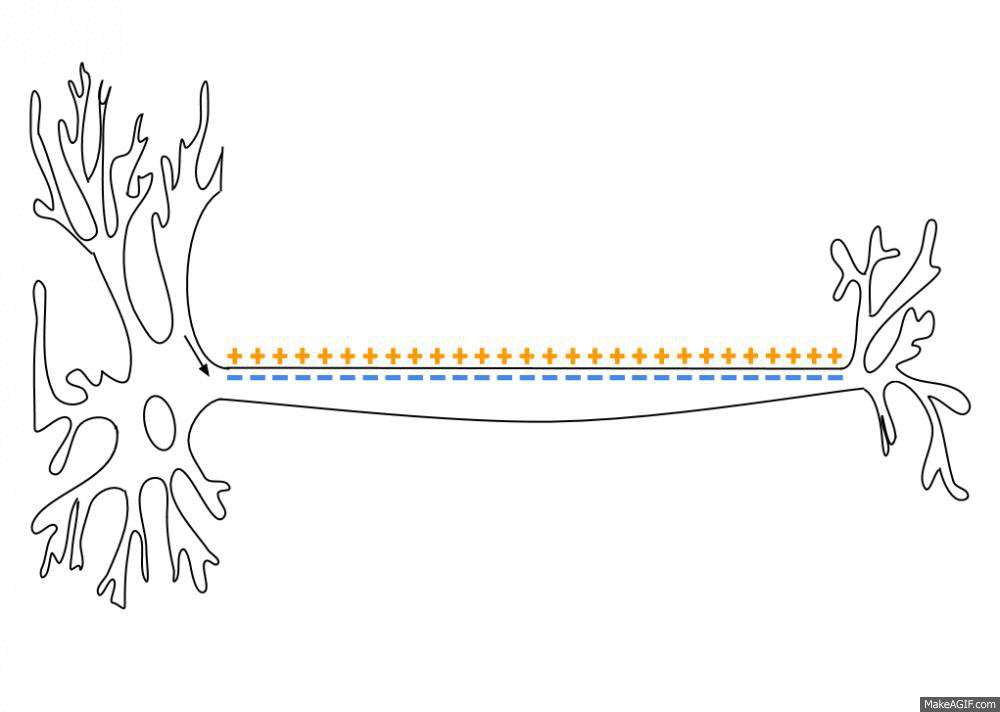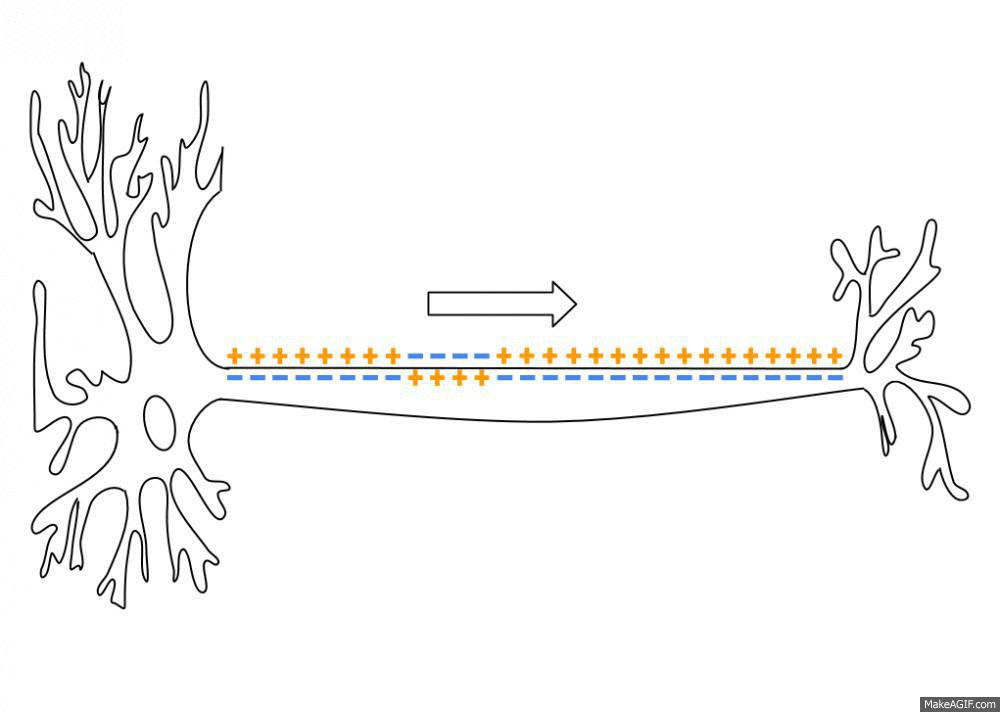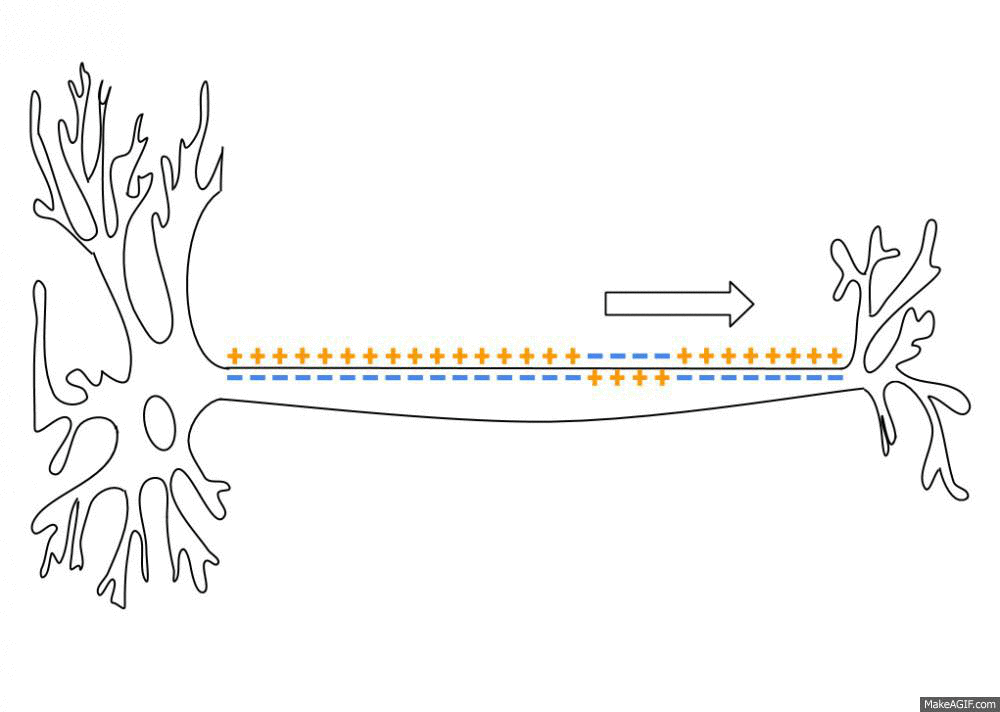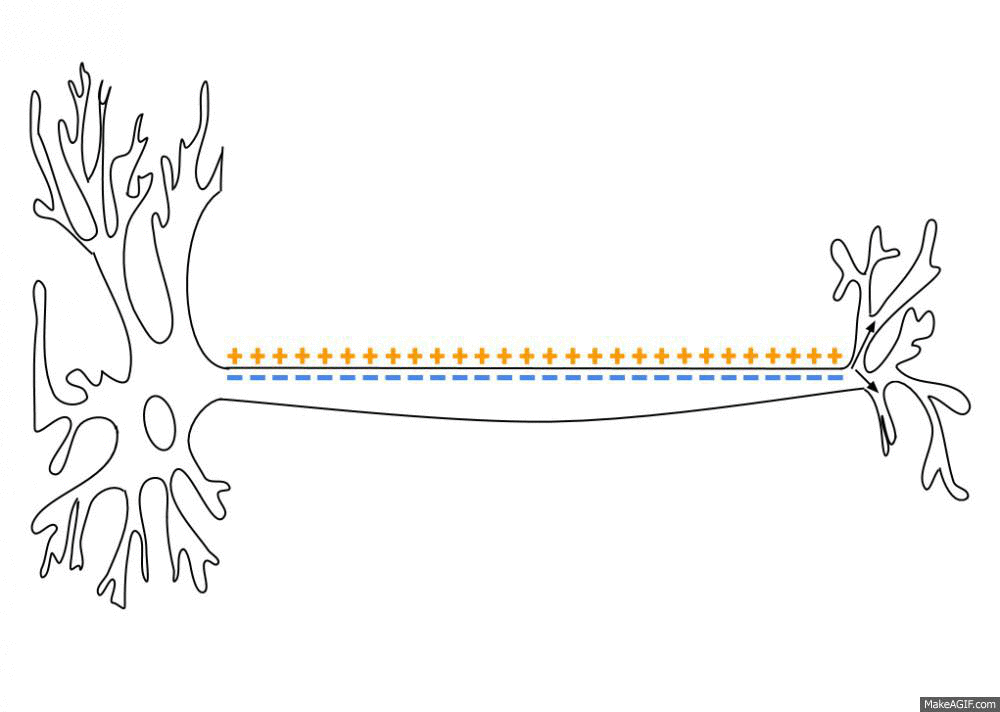
뉴런(neuron)은 축삭돌기(axon; 축삭)를 따라 전파되는 전기화학적 신호인 활동전위(action potential)를 발화하며 소통합니다. 이 신호는 축삭돌기 끝에서 신경전달물질(neurotransmitter)의 방출을 초래하여 신경계(nervous system)에 정보를 전송합니다. 활동전위는 급격한 전압의 상승을 초래하는 막전위(membrane potential)의 특정한 “실무율 (all-or-none)”변화입니다.
뉴런의 막전위
뉴런은 일반적으로 약 -70mV(밀리볼트)의 휴지막전위(resting membrane potential)를 가지고 있습니다. 예를 들어 신경전달물질이나 감각 자극으로부터 신호를 받으면 막전위는 자극의 특성에 따라 과분극(hyperpolarization; 더 음전하를 띰) 또는 탈분극(depolarization; 더 양전하를 띰)이 일어날 수 있습니다.
막이 특정 역치전위(threshold potential)까지 탈분극하면 전위의존성 나트륨 채널(voltage-gated sodium, 이하 Na+ 채널)이 반응하여 열립니다. Na+는 세포 내부보다 세포 외부의 농도가 높기 때문에 채널이 열리면 전기화학적 구배(electrochemical gradient)를 따라 세포 내부로 돌진합니다. 양전하가 유입되면서 막전위는 더욱 탈분극해 더 많은 채널을 엽니다. 그 결과, 막전위는 약 +40mV의 정상까지 빠르게 상승합니다.
활동전위가 최고조에 달했을 때, 몇 가지 요인이 전위를 다시 끌어내립니다. 일단 Na+ 채널이 비활성화하기 시작해 Na+의 유입이 느려집니다. 세포 내부가 더 양전하를 띨수록 Na+을 내부로 움직이는 전기 인력이 줄어듭니다. 초기 탈분극은 또한 전위의존성 칼륨 채널(voltage-gated potassium channel, 이하 K+ 채널)의 개방을 초래하지만 Na+ 채널보다 천천히 열립니다. 일단 이 K+ 채널이 활동전위의 정점 가까이 열리면 K+가 세포 밖으로 튀어나와 전기화학적 구배를 따라 내려갑니다. Na+를 통한 양전하 유입 감소와 K+에 따른 양전하 유출은 막전위를 빠르게 낮춥니다.
활동전위 후 짧은 기간 동안 막은 휴지전위(resting potential)에 비해 과분극됩니다. 이 기간을 불응기(refractory period)라고 부르며, 이때 세포는 새로운 활동전위를 생성할 수 없고, 따라서 세포에서 활동전위가 역으로 이동하는 것을 방지합니다.
전도성을 증가시키는 미엘린 수초(myelin sheath; 미엘린초)
특수 신경교세포(glial cell; 교세포; 신경아교세포), 즉 중추신경계의 희소돌기아교세포(oligodendrocyte)와 말초신경계의 슈반세포(Schwann cell)는 긴 돌기(process)를 연장하여 뉴런 축삭돌기를 감쌉니다. 이런 포장은 절연성을 제공해 전류가 축삭돌기를 따라 이동할 때 누출되는 것을 방지합니다. 또한, 전기 신호는 수초화(myelination; 미엘린화)된 영역에서 수동적인 양전류 흐름에 의해 수초화된 축삭돌기를 따라 전파됩니다. 전위의존성 Na+ 채널과 K+ 채널은 랑비에결절(node of Ranvier)에 있는 미엘린(myelin) 사이의 틈새에서만 볼 수 있으며, 이런 결절(node)에서 활동전위의 재생을 일으킵니다. 따라서 활동전위가 축삭돌기를 따라 이동할 때 결절과 결절 사이를 “점프”하는 것처럼 보이는데, 이를 도약전도라고 합니다(saltatory conduction).
오징어의 거대한 신경
동물학자이자 신경생리학자인 존 영(John Z. Young)은 오징어가 포유류 뉴런보다 훨씬 넓은 축삭돌기를 가진 신경세포를 가지고 있음을 발견했습니다. 이러한 신경은 더 큰 축삭돌기에서만 가능한 더 빠른 활동전위에 의해 촉진되는 빠른 탈출 기능을 조절합니다. 축삭돌기의 더 큰 안지름은 활동전위에 관련된 이온 기작에 대한 초기 연구와 설명을 가능하게 했습니다. 이런 지식은 1950년대 앨런 호지킨(Alan Hodgkin)과 앤드루 헉슬리(Andrew Huxley)가 대서양 오징어의 거대한 신경을 연구하면서 개척되었습니다. 그들은 축삭돌기 막의 나트륨 이온과 칼륨 이온에 대한 투과성을 설명했으며 전극 기록에 근거해 활동전위를 정량적으로 재구성할 수 있었습니다.