유전자의 수평적 전달
수평적 유전자 이동(水平的 遺傳子 移動, 영어: horizontal gene transfer, HGT 또는 lateral gene transfer, LGT)은 생식에 의하지 않고 개체에서 개체로 유전형질이 이동되는 현상을 가리키는 유전학의 개념이다. 수평적 유전자 전달(水平的 遺傳子 傳達) 또는 수평적 유전자 전이(水平的 遺傳子 轉移)라고도 한다. 주로 단세포 생물에서 관찰된다. 같은 종간에 이루어지는 박테리아 이동과는 달리 수평적 유전자 이동은 종간의 차이를 뛰어넘어 이동할 수 있다. 유전공학은 실험을 통해 인위적인 수평적 유전자 이동을 일으키기도 한다.
⑴ 바이러스
⑵ 박테리아의 재조합
⑶ 이동성 DNA
⑷ 원형질 연락사
바이러스

바이러스(Virus)는 살아있는 세포를 통해서만 생명활동을 하는 '존재'이다. 평상시에는 돌덩어리와 같은 상태로 비활성화된 상태로 있다가 살아있는 세포와 접촉하면 이에 기생하여 생명활동을 시작한다. 다른 세포가 가진 유전정보를 복제할 수 있는 능력은 있지만, 자체적으로 신진대사를 할 수는 없기 때문에 바이러스는 다른 생명체 없이 스스로 생명활동을 할 수 없다. 따라서 바이러스를 생명체라고 부를 수 있는지에 대해서는 논란이 있다. 이 때문에 생물 분류 단계에서 세균역, 진핵생물역, 고균역 그 어디에도 속하지 않는 미분류 상태이다. 현재는 생물과 무생물 사이 존재 정도로 취급되고 있다.
⑴ 바이러스(virus)
① 단위 : 플라크(용균반, plaque)
② 방출량(burst size) : 박테리아 내에 증식하는 파지의 숫자
③ 생물적 특성
○ 핵산과 단백질로 구성
○ 세포 내에서 활성을 나타낼 수 있어 자기 증식과 유전을 함
○ 돌연변이가 자주 일어나 환경에 빠르게 적응
④ 무생물적 특성
○ 단백질 결정체로 존재할 수 있음
○ 물질대사 효소 부존재 : DNA 복제 또는 전사와 관련된 효소는 존재
⑵ 구조
① 바리온(virion) : 바이러스의 기본 단위. 숙주밖에 있을 때 입자상태
② 핵산 + 캡시드(단위체 : 캡소미어) + 세포침투효소 + (세포막) + (역전사효소)
○ 세포막은 동물바이러스에만 존재
○ 역전사효소는 역전사 바이러스에만 존재 (예 : 레트로바이러스)
③ 꼬리 섬유 : 박테리오 파지 등에 존재. 숙주세포 표면에 존재하는 수용체를 특이적으로 인식하여 부착
⑶ 분류
① 외피의 유무 : 노출 바이러스, 외피 바이러스
○ 노출 바이러스(naked virus) : 외피 없음. 디네인으로 핵내 이동. 리소자임을 통한 세포 외 방출
○ 외피 바이러스(positive virus) : 외피 있음. 내막계를 통해 핵내 이동
② 숙주 : 박테리오 파지, 식물 바이러스, 동물 바이러스
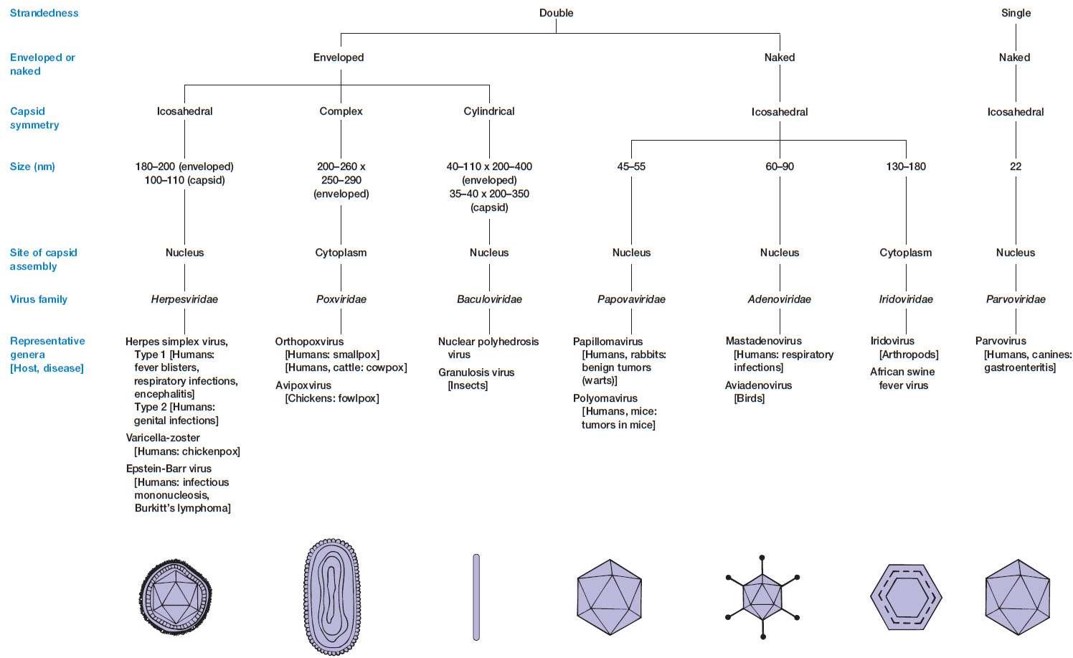
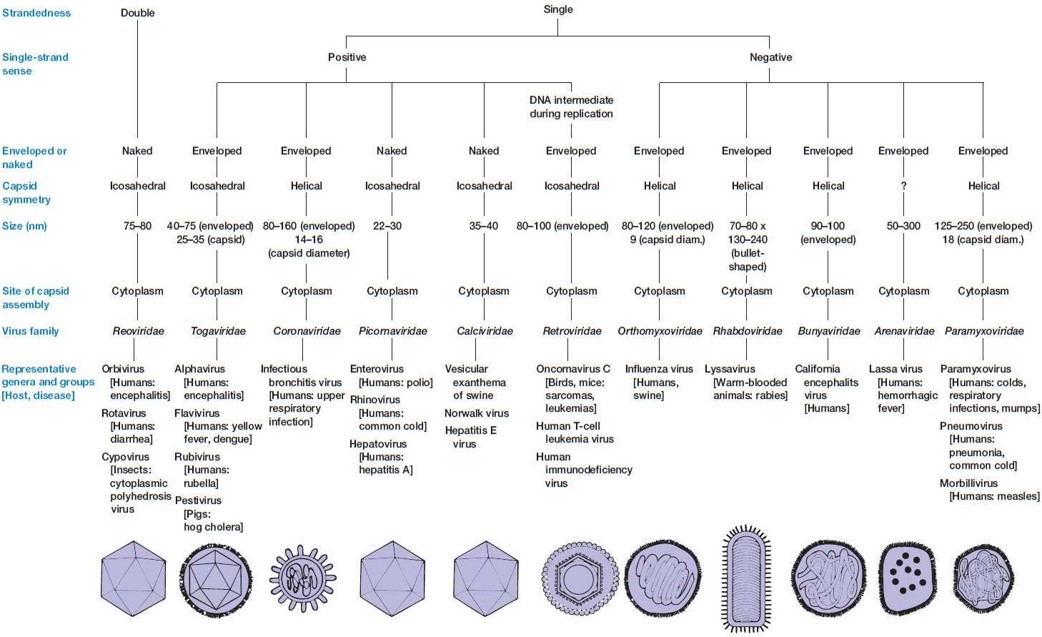
③ 유전물질에 따른 분류 : ds DNA, ss DNA, ds RNA, ss RNA
④ 당 인식 단백질에 따른 분류
○ 헤마글루티닌(hemagglutinin)
○ 숙주세포에 침투 시에 작용
○ 숙주세포 표면에 존재하는 탄수화물 말단에 존재하는 시알산 잔기들을 인식하여 결합
○ 시알산(sialic acid)은 N-아세틸뉴라민산(n-acetylneuraminic acid)의 별칭
○ 뉴라미니다아제(neuraminidase)
○ 숙주세포에서 방출 시에 작용
○ 동물 인플루엔자 바이러스 출아 시 숙주 원형질막의 시알산과 헤마글루티닌이 일시적으로 결합
○ 뉴라미다아제는 일시적 결합(글리코시드 결합)을 분해
○ 동물 인플루엔자 바이러스(influenza virus)는 헤마글루티닌과 뉴라미니다아제의 종류에 따라 세분화
○ 헤마글루티닌은 1번부터 16번까지 존재
○ 뉴라미니다아제는 1번부터 9번까지 존재
○ H#N#와 같은 형식으로 바이러스를 분류
○ 타미플루(Tamiflu™, oseltamivir) 및 릴렌자(Relenza™)
○ 뉴라미니다아제의 경쟁적 저해제. 시알산과 유사
○ 인플루엔자 바이러스 증식을 억제함
⑤ 항원변이기작에 따른 분류
○ 항원변이(antigenic drift) : 한 종류의 바이러스가 연속적 증식 → 돌연변이로 항원변이 생성
○ 항원대변이(antigenic shift) : 두 종류의 바이러스가 동일 숙주에서 증식 후 새로운 항원변이 생성, 바이러스의 분절된 유전체의 재편성 과정 有
○ 인플루엔자 바이러스는 항원대변이를 통해 다양성 획득
○ burst size : 박테리아 내에서 증식하는 파지의 숫자
⑷ 유전물질에 따른 분류 : ds DNA, ss DNA, dsRNA, ssRNA
① dsDNA virus
○ 인지질 이중층이 핵막에서 기원
○ 예 : AdV, HSV(Herpes virus), Hepatitis B virus, 천연두 바이러스(smallpox virus), VACV
② ssDNA virus
○ 인지질 이중층이 핵막에서 기원
○ 예 : H1-PV, Parvovirus
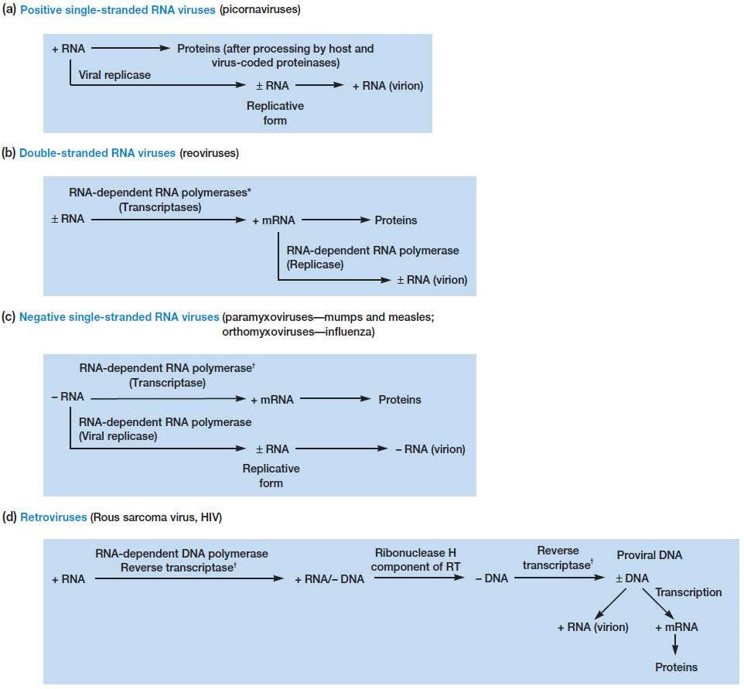
③ dsRNA virus : (+)는 번역, (-)는 RNA 합성의 주형(anti-sense)
○ 인지질 이중층이 원형질막에서 기원
○ 1st. 이중가닥 RNA가 (-) ssRNA와 (+) ssRNA로 분리
○ 2nd. (-) ssRNA는 자신의 효소 (RNA dependent RNA pol)을 사용하여 dsRNA 생성
○ 3rd. (+) ssRNA는 mRNA로 기능하여 바이러스 단백질 번역
○ 예 : reovirus, rotavirus
④ ssRNA virus : (-) negative-sense ssRNA virus, (+) positive-sense ssRNA virus, 레트로바이러스로 분류
○ 인지질 이중층이 원형질막에서 기원
⑤ (+) positive-sense ssRNA virus : (+)는 번역, (-)는 RNA 합성의 주형(anti-sense)
○ positive strand : 기본적인 유전정보를 가지고 있음
○ mRNA로 기능하여 바이러스 단백질을 번역해야 하므로 5' cap과 poly A 꼬리가 존재
○ 1st. 자신의 RNA pol을 사용하여 (-) ssRNA 다수 합성
○ 2nd. (-) ssRNA는 자신의 효소(RNA dependent RNA pol)를 이용하여 (+) ssRNA 다수 합성
○ 3rd. (+) ssRNA는 유전체 구성 및 단백질 번역에 이용
○ 유전체로 구성할 RNA가 번역에도 사용되므로 돌연변이율 높음
○ 예 : Hepatitis C virus, 사스(SARS), 코로나 바이러스(Corona virus), poliovirus, coxsackievirus, NDV
⑥ (-) negative-sense ssRNA virus : (+)는 번역, (-)는 RNA 합성의 주형(anti-sense)
○ negative strand : positive strand의 보완. negative strand의 유전정보를 실제로 쓰지는 않고 복제 주형으로만 사용
○ 1st. 자신의 RNA dependent RNA pol을 이용하여 (+) ssRNA 다수 합성
○ 2nd. (+) ssRNA는 자신의 효소(RNA dependent RNA pol)을 이용하여 (-) ssRNA 다수 합성
○ 3rd. (-) ssRNA는 유전체 구성에 이용, (+) ssRNA는 단백질 번역에 이용
○ 예 : 인플루엔자 바이러스, 에볼라 바이러스, MV, NDV, VSV
○ 암기팁. 인플루엔자.. 음플루엔자..
⑦ 레트로바이러스(retrovirus) : 역전사 효소(RT, reverse transcriptase)가 존재, (+) positive-sense ssRNA virus의 일종
○ 역전사 바이러스 : 역전사 효소(RT, reverse transcriptase)가 존재하는 바이러스
○ 역전사 바이러스는 레트로바이러스와 비 레트로바이러스(예 : dsDNA virus인 Hepatitis B virus)로 분류
○ HIV virus
○ 동일한 ssRNA 가닥 두 개가 유전체를 구성. 상보적인 가닥이 아님을 유의
○ HIV RNA는 5'-cap과 poly-A tail이 있음
○ HIV virus 내 주요 효소 : 역전사 효소(RT), 삽입효소(integrase), 단백질 분해효소(protease)
○ (참고) 인플루엔자 바이러스의 유전체는 RNA이고 이 RNA로부터 RNA를 바로 복제할 수 있음
○ 단, 조류독감 인플루엔자의 경우 cDNA를 합성하는 역전사 과정이 존재
⑸ 박테리오 파지 : 생활사 주기가 약 30분
① 용균성 생활사(lytic cycle) : T1, T2, T4 파지 등 병원성 바이러스(DNA virus)
○ 1st. 부착 : 숙주의 세포 표면에 꼬리 부착
○ 2nd. 침투 : 파지의 작용으로 숙주의 복제, 전사, 번역이 즉시 중단
○ 3rd. 복제 : 초기 유전자가 발현 → 후기 유전자 전사 유도 → 후기 유전자 발현
○ 초기 유전자 : 생활사 결정 유전자, 숙주 DNA 파괴 유전자(숙주 염색체를 모두 가수분해시켜 자신 DNA 합성에 이용)
○ 후기 유전자 : 캡시드 유전자, 용해 유전자
○ 4th 조립 : 숙주 효소로 바이러스 DNA 복제. 생산된 바이러스 단백질은 숙주 DNA를 절단 및 분해함
○ 5th. 방출 : 파지가 생산한 세포막 분해효소에 의해 방출
② 용원성 생활사(lysogenic cycle) : λ 파지 등 온건성 파지
○ 1st. 부착
○ 2nd. 침투 : DNA가 박테리아 내로 삽입되면 선형 DNA가 환형 DNA로 전환
○ 3rd. 복제 : 용균성 생활사와 달리 파지 DNA가 숙주 DNA에 삽입
○ 프로파지 : 숙주 DNA에 삽입된 파지 DNA
○ 4th. 조립 및 방출 : 원래 파지 DNA는 숙주에 계속 남아 있음
○ 5th. 숙주가 증식하지 못하면 용균성 생활사로 전환
○ cI 전사인자가 많으면 용원생 생활사, cro 전사인자가 많으면 용균성 생활사
○ 암기팁. I.. in.. O.. out..
○ 건강한 대장균에서 파지는 처음에 cro를 합성하다가 나중에 합성되는 cI 단백질의 영향으로 용원성 생활사를 함
③ cro와 cⅠ의 경쟁적 발현
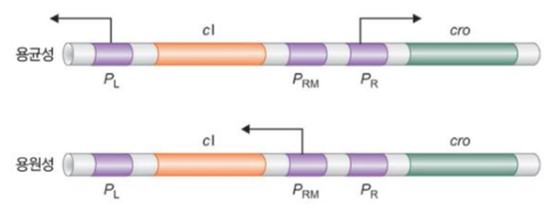
○ 용균성 생활사와 용원성 생활사는 두 개의 유전자(cⅠ, cro)와 세 개의 프로모터(PR, PL, PRM)로 조절됨
○ 용균성 생활사에서의 유전자 발현 양상
○ 1st. cro로부터 생성된 단백질이 PRM 내에 있는 OR3에 결합하여 cⅠ의 전사를 억제
○ 2nd. 그 결과 용균성 성장을 결정하는 시기에 PL과 PR 프로모터가 활성화됨
○ 용원성 생활사에서의 유전자 발현 양상
○ 1st. cⅠ 유전자에 의해 λ 억제인자가 생성됨
○ 2nd. λ 억제인자는 PR 내에 있는 OR1과 PRM과 PR에 걸쳐있는 OR2에 결합하여 cⅠ의 발현을 활성화
○ 3rd. 대신 cro의 발현이 억제됨
○ 4rh. 그 결과 용원성 성장을 결정하는 시기에 PRM 프로모터가 활성화됨
④ 용원성 유도
○ 1st. 용원성 유도
○ 1st - 1st. 성장조건이 좋지 않은 배지에서 cⅡ 단백질의 합성이 증가
○ 1st - 2nd. cⅡ 단백질은 PRE라는 프로모터의 상단에 결합
○ 1st - 3rd. OR1 - OL1, OR2 - OL2, OR3 - OL3 간에 억제인자를 사이에 두고 DNA 고리를 형성
○ 1st - 4th. DNA 고리는 cⅠ 유전자의 전사를 촉진
○ 1st - 5th. cro와 cⅠ의 경쟁적 유전자 발현에 의해 cⅠ의 발현이 증가함에 따라 용원성 생활사가 유도
○ 2nd. 용원성 확립
○ cⅠ 유전자가 용원성을 확립하는 경우 PRE에서 전사되고, 그 상태를 유지할 경우 PRM에서 전사
○ OR1과 OR2에 결합한 λ 억제인자도 용원성 확립을 활성화
○ PR과 cⅡ도 용원성 확립에 영향을 줌
○ 용균성 유전자를 조절하는 PL도 용원성 확립에 영향을 줌
⑤ 용균성 유도
○ λ 억제인자는 N 말단과 C 말단을 양 기점으로 아령 모양으로 생김
○ λ 억제인자 단량체는 2량체를 형성하고, 2량체가 서로 결합하여 4량체를 형성함
○ UV는 λ 억제인자가 단량체가 되도록 하여 cⅠ의 발현을 억제
○ cro와 cⅠ의 경쟁적 유전자 발현에 의해 cro의 발현이 상대적으로 증가함에 따라 용균성 생활사가 유도됨
⑥ 항종결(antitermination) : 람다파지의 N과 Q 단백질과 관련
⑦ 역조절(retroregulation) : int 발현에 있어 헤어핀 구조가 핵산분해효소에 저항성을 가짐
⑹ 동물 바이러스
① 동물 바이러스의 침투방식
○ 노출바이러스 : 바리온이 엔도사이토시스로 흡수된 뒤 소낭의 막을 파괴
○ 외피 바이러스
○ 첫 번째 방식 : 엔도사이토시스로 흡수된 후 외피와 소낭의 막이 융합
○ 두 번째 방식 : 숙주세포의 원형질막과 바이러스의 외피가 융합
② 인플루엔자 바이러스 : ssRNA 바이러스
○ 총 8개의 RNA 분절로 구성됨
○ 캡시드에 직접 RNA 중합효소가 붙어 있음
○ 인플루엔자의 분류 : 헤마글루티닌과 뉴라미다아제의 종류에 따라 분류
○ 1st. 바이러스 외피막의 당단백질 스파이크가 세포막의 수용체와 결합
○ 2nd. 외피와 소낭의 막 융합 → 세포질로 바리온 방출
○ 3rd. RNA-dependent RNA polymerase 활성
○ 4th. 바이러스 단백질 합성 → budding
③ HIV : ssRNA 바이러스
○ 캡시드 내 reverse transcriptase, protease, integrase, 동일(↔ 상보성) ssRNA × 2개 존재
○ 1st. 외피에 나와 있는 당단백질 gp 120이 보조 T세포의 CD4와 결합
○ 피막은 당단백질 gp 120과 당단백질 gp 41로 구성
○ 당단백질 gp 120은 CD4를 인식
○ 당단백질 gp 41은 CCR5를 인식
○ CCR5가 없는 개체는 HIV 저항성이 나타남
○ 2nd. 원형질막과 융합하여 세포 내로 침투하고 효소에 의하여 캡시드 제거
○ (참고) 캡시드 P24 : HIV 바이러스 감염을 진단하는 1순위 마커
○ (참고) 바이러스 감염을 진단하는 2순위 마커는 항체
○ 3rd. 역전사 효소가 작용
○ 3rd-1st. RNA-dependent DNA polymerase 활성 : poly T를 프라이머로 mRNA 역전사. ssRNA → ss cDNA
○ 3rd-2nd. RNAaseH 활성
○ 3rd-3rd. DNA-dependent DNA polymerase 활성 : DNA pol Ⅰ 기능. ss cDNA → ds cDNA
○ 4th. 이중나선 DNA(ds cDNA)는 프로바이러스가 되어 핵에서 잠복
○ 5th. RNA 전사 : 대체 스플라이싱 후 캡시드와 핵산 형성
○ 6th. 바이러스가 원형질막으로 둘러싸인 후 빠져 나감
⑺ 식물 바이러스
① 수평 감염 : 원형질 연락사를 통해 인접 세포로 바이러스가 이동
② 수직 감염 : 번식 과정과 함께 세대를 거쳐 식물 바이러스가 감염되는 형태
⑻ 주요 바이러스 질환
① HBV(hepatitis B virus) : 간암, IFG-2(인슐린유사 성장인자)의 발현 증가
② SV40(simian virus40) : 원숭이 림프종, SV40의 T-항원에 의해 pRb, p53의 억제
③ Herpes simplex virus(포진유발) : 림프종, 카포시 육종 등, 종양세포의 MHC I에 항원제시 억제
④ HTLV(human T-leukemia virus) : T세포 림프종, T세포에 감염되어 사이토카인의 과다분비
⑤ HIV 바이러스