광합성
光合,光合作用, Photosynthesis
광합성은 생물이 빛을 이용하여 양분을 스스로 만드는 과정으로, 물과 이산화탄소를 재료로 포도당과 산소를 생성한다.

암반응 (캘빈 회로
⑴ 개요
① 광합성 생물 : 박테리아, 조류, 식물에 해당
○ 종류 1. 박테리아 : 홍세균, 황세균, 남세균
○ 종류 1-1. 홍세균
○ 870 nm 파장 흡수. 광계 1개
○ 순환적 광인산화 only
○ H2S가 전자 공여
○ 역전자전달 (∵ 집광을 하지 못함)
○ 종류 1-2. 황세균
○ 840 nm 파장 흡수. 광계 2개
○ 순환적 광인산화 + 원시적 비순환적 광인산화
○ H2S가 전자공여
○ 종류 1-3. 남세균 : 엽록체의 조상
○ 670 nm 파장 흡수. 광계 2개
○ 순환적 광인산화 + 비순환적 광인산화
○ H2O가 전자공여
○ 종류 2. 조류
○ 홍조류 : 엽록소 a, d
○ 규조류 : 엽록소 a, c
○ 갈조류 : 엽록소 a, c
○ 녹조류 : 엽록소 a, b
○ 종류 3. 선태식물, 양치식물, 겉씨식물, 속씨식물
○ 선태식물 : 물질교환을 확산으로 함, 엽록소 a, b
○ 양치식물, 겉씨식물, 속씨식물 : 엽록소 a, b
○ 음지식물 : 카로티노이드계 색소가 없음
② 반응식 : 6 CO2 + 12 H2O + 빛 → 포도당(C6H12O6)+ 6O2 + 6H2O (ΔG = 686 kcal / mol)
○ 세포호흡의 반대 결과를 달성
○ 광합성 반응식의 추적

○ 닐의 연구 : 황세균을 이용하여 광합성 산물인 산소의 기원이 물임을 밝혀냄
○ 루벤의 연구 : 방사성 동위원소 18O를 이용하여 닐의 가설을 증명
③ 광합성 실험
○ 엥겔만(Engelmann) 실험
○ 해캄과 호기성 세균을 이용한 광합성 효율 실험
○ 엽록소는 400 ~ 500 nm 파장과 600 ~ 700 nm 파장의 빛을 잘 흡수하나 500 ~ 600 nm 파장의 빛은 아님
○ 즉, 청자색과 적황색 파장의 빛에서 광합성이 효율적으로 일어남
○ Emerson experiment - Emerson red drop
○ 파장 대 양자 수율에 대한 그래프를 제시
○ 양자수율 : 흡수 스펙트럼과 광합성 효율에 대한 함수
○ 흡수 스펙트럼이 680 nm에서 피크이고 700 nm 이후 급감 : 광계 Ⅱ가 작동하지 않아 광합성 효율이 감소해서
○ Emerson experiment - Emerson enhancement effect
○ Duysens' experiment
○ 벤슨의 실험
④ 광합성 원리
○ CO2와 O2는 기공을 통해서 운반
○ 명반응(광의존성 반응)
○ 빛 에너지를 이용하여 전자전달
○ 환원반응 : NADP+ → NADPH, ADP → ATP
○ 물 분해, 산소 방출
○ 암반응(광비의존성 반응)
○ CO2를 환원하여 C6H12O6을 생성
○ 산화반응 : NADPH → NADP+, ATP → ADP
⑤ 광합성 요인
○ 최소량의 법칙
○ 식물의 성장은 넘치는 것이 아니라 가장 부족한 필수 영양소에 의해 좌우된다는 법칙
○ 1840년 독일의 식물학자 유스투스 리비히가 주장
○ 종속변인 : CO2
○ 조작변인 1. 빛 : 광포화점 有

○ 순광합성량 : 외관상 광합성량
○ 총광합성량 : 실제 광합성에 사용된 모든 CO2 양
○ 호흡량 = 총광합성량 - 순광합성량
○ 보상점(B) : 총광합성량과 호흡량이 같아질 때의 빛의 세기
○ 광포화점(D) : 광합성량이 최대가 되기 시작하는 빛의 세기
○ 양지식물은 더 크고 물질대사가 활발하므로 음지식물보다 광포화점과 보상점이 높음
○ 양지식물은 카로티노이드가 있어 광포화점이 큼
○ 조작변인 2. 온도 : 최적온도 有
⑵ 엽록체
엽록체(葉綠體, chloroplast)는 진핵생물에서 광합성을 하는 세포 소기관
⑶ 색소
① 광합성 색소 분리 실험 : 종이 크로마토그래피 (톨루엔 용매)
② 엽록소 : 엽록소 a와 엽록소 b로 구분, 적색광과 청색광 흡수
○ 포르피린 고리
○ 방향족성이 있어 색소로 기능
○ 구조 : 마그네슘 보결족 + 테트라피롤(pyrrole) + 피톨
○ 탄화수소 꼬리(피톨) : 엽록체의 틸라코이드막 내에 위치하는 단백질(광계 등)의 소수성 부분과 상호작용
○ 엽록소 a : 메틸기
○ 엽록소 b : 알데히드기
③ 카로티노이드 : 보조색소. 녹색광 흡수. 엽록소보다 광합성 효율은 떨어짐. 양지식물에게만 존재
○ 기능 1. 스펙트럼 확장
○ 기능 2. 강한 빛으로부터 광합성 기구 보호
○ β-카로틴 : 카로티노이드의 한 종류
○ 400 ~ 500 nm의 빛을 주로 흡수하지만 녹색광도 흡수
○ 비타민 A로 전환 가능
○ 비타민 A의 유도체인 레티날은 망막의 간상체와 추상체에서 시각 색소의 구성 성분으로 이용
④ 크산토필
⑤ 피코빌린 : 홍세균(870 nm), 황세균(840 nm), 남세균(670 nm)의 색소 분자
⑷ 광계
① 반응중심 : 틸라코이드막에서 빛에너지가 화학에너지로 바뀌는 장소
○ 엽록소 a 2개, 1차 전자수용체, 단백질 등으로 구성
○ 광계 Ⅰ : 700 nm 파장을 흡수. 스트로마에 노출되어 역사적으로 먼저 발견
○ 광계 Ⅱ : 680 nm의 파장을 흡수. 스트로마로부터 가려져 역사적으로 늦게 발견
○ 엽록소 a가 충분한 에너지 흡수 → 들뜬 전자의 궤도 이탈 → 고에너지 전자는 전자전달계로 전달
② 집광복합체(안테나) : 반응중심 주변부, 엽록소 a, 엽록소 b, 카로티노이드, 크산토필 등 존재
⑸ 명반응 : 엽록소 분자로 덮인 원반형 막 구조인 틸라코이드(thylakoid) 막에서 일어난다.
① 1st. 안테나 : 광자는 색소 분자에 흡수되고 RET(resonance energy transfer)에 의해 인접 색소분자로 광자 이동
○ RET : 두 분자가 가까이 붙어 있으면 형광 기작이 아닌 보다 직접적인 방법으로 광자가 전달되는 현상
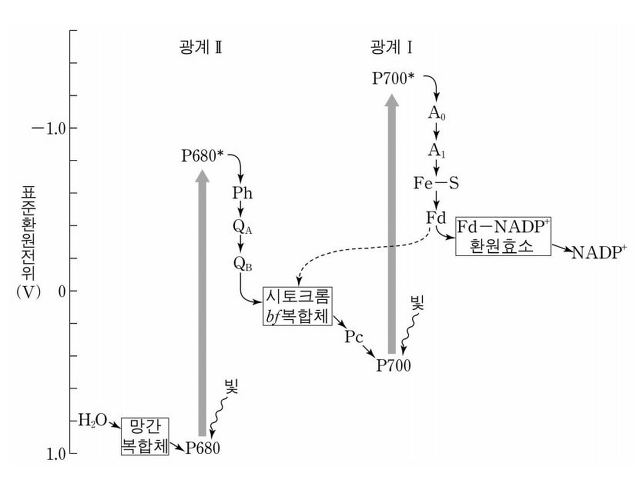
② 2nd. 반응중심 : 전자는 색소 분자로 옮겨 다니다가 광계 Ⅱ의 P680 또는 광계 Ⅰ의 P700으로 이동
○ P680은 680 nm에서 최대 흡광도를, P700은 700 nm에서 최대 흡광도를 보임
③ 3rd. 광계 Ⅱ의 P680이 P680+로 산화돼 1차 전자 수용체(primary acceptor)로 전달
○ 1차 전자 수용체를 페오피틴(pheophytin)이라고도 함
○ P680+은 지구상에서 가장 강한 산화제
④ 4th. P680은 페오피틴의 중심에 Mn이 있는 구조로, 산화된 P680+는 H2O을 산화시켜 전자를 획득 (O2 생성)
○ 물의 광분해는 틸라코이드 내부에서 일어남
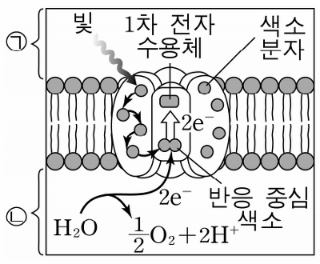
⑤ 5th. 1차 전자 수용체로 이동한 전자는 PQ(plastoquinone), ATP 합성효소(cytochrome b0f complex, CF0CF1 complex), PC(plasto cyanine)를 거쳐 P700에 도달
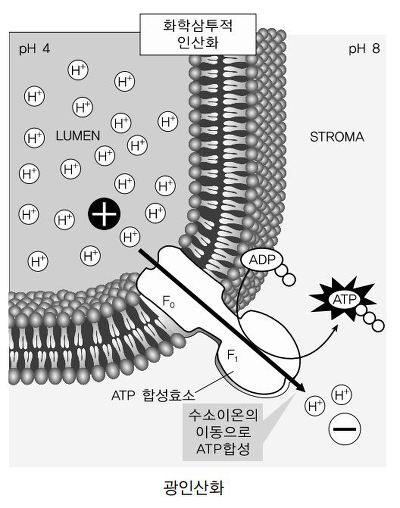
○ 이 과정에서 3H+가 이동하여 1 ATP 생성
○ ATP 합성효소는 양성자의 촉진확산을 수행하는 CF0 채널과 ATP 합성효소를 포함하는 CF1으로 구성되어 미토콘드리아의 F0F1과 같은 화학삼투 인산화 방법으로 ATP 생성
⑥ 6th. ②와 ⑤에 의해 전자를 받은 P700은 들뜬 상태인 P700*가 됨, 그 뒤 전자는 1차 전자 수용체(primary accepter)로 전달
○ 1차 전자 수용체를 필로퀴논(phylloquinone)이라고도 함
○ P700*는 세상에서 가장 강한 환원제
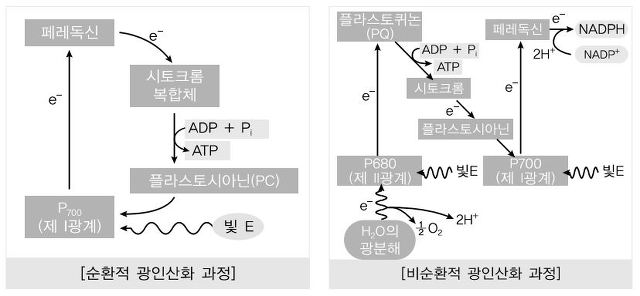
⑦ 7th. 전자가 필로퀴논에서 페레독신(ferredoxin)으로 전달
⑧ 8th - 1st. 순환적 광인산화 : 전자가 페레독신에서 시토크롬 복합체로 이동하는 경우. 1 ATP 생성
○ 산물 : ATP
⑨ 8th - 2nd. 비순환적 광인산화 : 페레독신은 ferredoxin-NADP+로 전자 전달
○ ferredoxin-NADP+의 환원작용으로 인해 1 ATP + 1 NADPH 생성
○ 산물 : ATP, NADPH, O2
○ NADP+가 최종 전자수용체
⑨ 억제제
○ 광계 Ⅰ 억제제 : 파라쿼트
○ 광계 Ⅱ 억제제 : DCMU
⑹ 암반응 (캘빈 회로, Calvin Cycle)
① 스트로마 : 엽록체의 기질. 암반응 장소
○ 포도당으로 설탕을 합성하는 반응은 세포질에서 일어남
② 캘빈회로(Calvin Cycle) : ATP와 NADPH를 사용하여 CO2를 환원하여 탄수화물 합성 (12 NADPH, 18 ATP)
○ 포도당 신생합성 과정의 일부를 진행 : 3PGA → 1,3-BPG → G3P
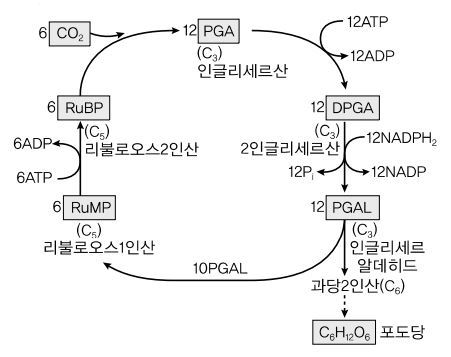
③ Rubisco(ribulose-1,5-bisphosphate carboxylase oxygenase)
○ 잎 전체 단백질의 50%, 지구상에 가장 풍부한 단백질
○ 이산화탄소↑ → 카르복실화 효소 작용, 산소↑ → 산소화 효소 작용
④ G3P : 스트로마에서 녹말이 되거나 엽육세포 세포질에서 sugar가 됨
⑤ 암반응 효소들은 광조건에서 더 효율적으로 기능
○ Rubisco
○광조건 → 스트로마 알칼리화 → 루비스코 활성 증가
○ 명반응으로 인한 pH 변화가 틸라코이드 내강에 저장된 Mg2+를 스트로마로 이동 → Rubisco 활성화
⑺ C3, C4 및 CAM 식물
① C3 식물 : 일반적인 식물(예 : 장미, 벼 등 대부분의 온대식물)
○ 최초 탄소고정 산물 : 3-PGA (3C)
○ 높은 온도에서의 반응
○ 공변세포 : 기공의 개폐 조절과 증산작용
○ 열린 기공 : 이산화탄소 확보 가능, 물 손실
○ 온도↑ → 기공닫힘 → CO2 부족 → 광합성률 감소
○ 온도↑ → 기공 닫힘 → CO2 부족 → 산소과다로 광호흡 발생
② 광호흡(Photorespiration)
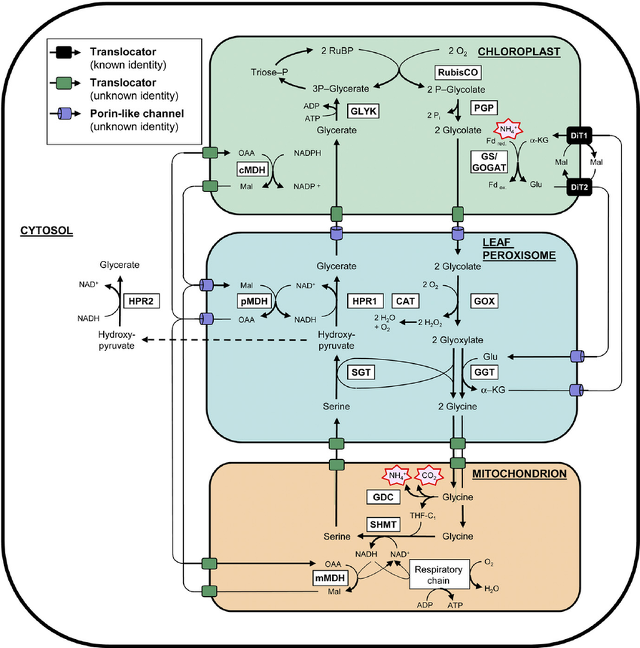
○ 루비스코는 이산화탄소를 기질로 할 수도 있고, 산소를 기질로 할 수도 있음
○ Rubisco가 산소화효소로 작용
○ 1st. RuBP + O2 → 3PG + 글리콜산(2C)
○ 2nd. 3PG + 글리콜산(2C) + ATP → RuBP + CO2
○ 글리콜산은 분해되어 탄소원을 잃어버림
○ 광호흡 과정에서 엽록체, 퍼옥시좀, 미토콘드리아가 관여
○ 광호흡은 에너지를 사용하므로, 광호흡 환경(높은 온도)에서 순환적 광인산화 우세
○ 글리신, 세린이 중간체로 관여
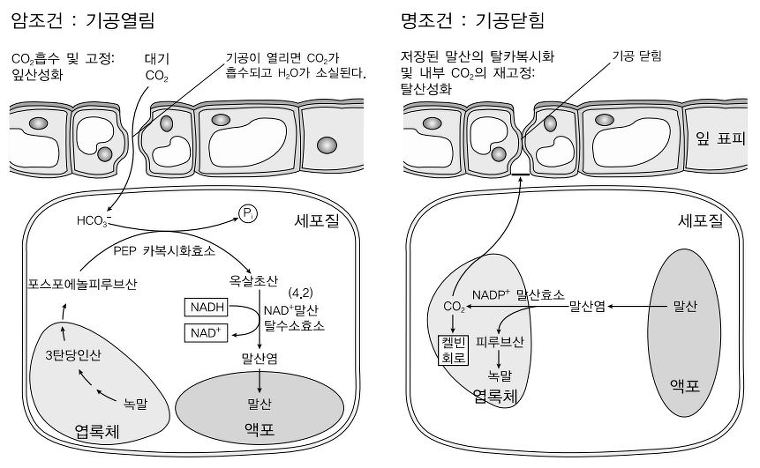
③ C4 식물 : 열대·아열대 식물(예 : 옥수수, 사탕수수, 수수 등 열대식물)
○ 광계 Ⅱ가 없음 : 정확하게는 엽육세포는 광계 Ⅰ, Ⅱ가 있으나 유관속 초 세포는 광계 Ⅰ만 있음 (ref)
○ 오직 순환적 광인산화, 보상점 낮음, 탄소고정이 공간적으로 분리
○ 크란츠 구조(Krantz anatomy) : 유관속 초 세포가 유관속을 둘러싸고, 엽육세포가 다시 그 주위를 둘러싸는 구조
○ 1st. 기공이 거의 닫힌 상태, 기공 인접 세포(엽육세포, mesophyll)에서 CO2에 친화력이 큰 PEP에 탄소고정
○ C4 식물의 엽육세포는 엽록체가 없음
○ 2nd. PEP 카르복실화효소가 PEP(3C)에 CO2를 결합 → 옥살로아세트산(OAA, 4C) 생산. 최초 탄소고정 산물
○ PEP 카르복실화효소는 루비스코보다 이산화탄소 친화력이 높음
○ PEP 카르복실화효소는 유관속 초 세포에 존재하지 않음
○ PEP 카르복실화효소(PEP carboxykinase)
○ 3rd. 옥살로아세트산이 환원된 말산(4C)은 심부(유관속 초 세포, bundle-sheath cell)로 이동 후 CO2 방출
○ 4th. 유관속 초 세포 내 Rubisco는 공급받은 이산화탄소 및 스트로마 내 옥살로아세트산과 함께 켈빈회로
○ Rubisco는 엽육세포에 존재하지 않음
○ 5th. CO2를 방출한 말산은 다시 NADPH를 생성하며 피루브산(3C) 생성 : 광계 Ⅱ가 없어도 되는 이유
○ 6th. 피루브산은 ATP를 사용하여 PEP 생성
○ 7th. PEP는 엽육세포로 이동
○ 다른 경로 : 옥살로아세트산이 말산이 아니라 아스파르트산을 경유
④ CAM 식물 : 초고온·건조 환경의 식물, 다육식물(예 : 돌나무, 선인장, 파인애플 등 사막식물)
○ 광계 Ⅱ가 있으나 광호흡이 잘 일어나는 조건에서 광계 Ⅱ의 활성이 감소함 (ref)
○ 오직 순환적 광인산화, 보상점 낮음, 탄소고정이 시간적으로 분리
○ 1st. 낮에는 기공을 완전히 닫아 수분 보존, 암반응도 일어나나 효율이 떨어짐
○ 2nd. 밤에 기공을 열어 C4 회로에서 CO2를 말산으로 생산
○ 2nd - 1st. PEP 카르복실화효소가 PEP에 CO2를 결합하여 옥살로아세트산 생산. 최초 탄소고정 산물
○ 2nd - 2nd. 옥살로아세트산은 말산으로 환원
○ PEP 카르복실화효소(PEP carboxykinase)
○ 3rd. 생산된 말산은 액포에 저장
○ 4th. 낮 동안 말산을 분해하여 얻은 CO2로 캘빈 회로 작동
○ 4th - 1st. 말산 → CO2 + NADPH + 피루브산(3C)
○ 4th - 2nd. 피루브산(3C)은 ATP를 사용하여 PEP가 되거나 TCA 회로에 사용되어 ATP 생산
○ 4th - 3rd. CO2는 스트로마로 이동하여 옥살로아세트산 및 루비스코와 함께 캘빈회로
⑤ C3, C4, CAM 식물의 비교
| C3 식물 | C4 식물 | CAM 식물 | |
| 1차 CO2 수용체 | RuBP | PEP | PEP |
| CO2 고정효소 | rubisco | PEP 카르복실화 효소 | PEP 카르복실화효소 |
| 최소 CO2 산물 | 3PGA | OAA | OAA |
| 최초과정 | CO2 + H2O + RuBP → 2PGA | CO2 + H2O + PEP → OAA | |
| 명반응 | 엽육세포 | 엽육세포 | 엽육세포 |
| 켈빈-벤슨회로 | 엽육세포 | 유관속초세포 | 엽육세포 |
| CO2 고정장소 | 엽육세포 | 엽육세포 | 엽육세포 |
| CO2 고정시간 | 낮 | 낮 | 밤 |
| 광호흡 | 강함 | 약함 | 약함 |
| 1g 광합성 시 필요 물의 양 | 약 500 g | 약 300 g | 약 50 g |
C3, C4, CAM 식물의 비교
⑻ 지구온난화와 광합성
① 온도 상승은 C3, C4, CAM 식물의 상대적 비율 및 분포에 영향
② 이산화탄소는 현재 광합성이 제거할 수 있는 것보다 더 빠르게 방출
③ 대기에 추가되는 CO2의 25%는 열대 삼림의 남벌(벌목과 화재)이 원인
④ 식목은 도움이 됨; 어린 나무는 늙은 나무보다 순광합성 속도가 빠름